№ 3 (18) 2018
Размерно-массовые характеристики слоевища
и ценопопуляции Dictyota fasciola (Roth) J.V. Lamouroux (Ochrophyta) и их изменчивость в Черном море
Dimension-mass properties of the thallus and cоenopopulations
of the Dictyota fasciola (Roth) J.V. Lamouroux (Ochrophyta) and their variability in the Black Sea
Евстигнеева И.К., Танковская И.Н.
Irina К. Evstigneeva, Irina N. Tankovskaya
Институт морских биологических исследований им. А.О.Ковалевского РАН
(Севастополь, Россия)
УДК 582.275.54+581.55
Впервые в ходе многолетних исследований были определены размерно-массовые параметры слоевища и ценопопуляций черноморской макроводоросли Dictyota fasciola (Roth) J.V. Lamouroux (Ochrophyta). Установлены их крайние, средние значения, пространственно-временная локализация и изменчивость. Выявлены константные и динамичные признаки слоевища и ценопопуляции.
Ключевые слова: Dictyota fasciola (Roth) J.V. Lamouroux; Ochrophyta; слоевище; ценопопуляция; размерно-массовый состав; динамика; Крым; Черное море.
Введение
В последнее время наблюдается ухудшение экологических условий на мелководных участках Черного моря, где сосредоточены основные заросли водорослей – продуцентов органического вещества и кислорода, кормовая база и убежище для многих животных. Водоросли чутко реагируют на изменения в окружающей среде и концентрируют в своих талломах многие химические элементы. Поэтому они могут быть не только индикаторами разных типов загрязнения среды, но и своего рода фильтрами, очищающими ее. Это делает актуальным изучение эколого-биологических особенностей олигосапробных видов водорослей, количественные и качественные характеристики которых могли бы адекватно отражать условия среды.
К таким видам относится Dictyota fasciola (Roth) J.V. Lamouroux – малоизученная черноморская водоросль, входящая в состав прибрежных цистозировых фитоценозов в качестве субдоминанта и в отдельные периоды года играющая заметную роль среди донной растительности (Танковская, Евстигнеева, 2002). Она относится к отделу Ochrophyta, порядку Dictyotales Kjellm, семейству Dictyotaceae J.V. Lamour. ex Dumort., роду Dictyota J.V. Lamour. (Guiry, Guiry, 2016). Слоевище этого вида пластинчатое, грубокожистое, плоское, дихотомически правильно разветвленное на узкие ветви-сегменты, прикрепляется подошвой или стелющимися узкими разветвленными столонами, часто по краю прорастает, с беспорядочно отходящими проростками различной длины (Зинова, 1967). Вид поселяется на каменистом грунте вулканического или осадочного происхождения в исключительно чистых местах открытой части моря. В Черном море он получил существенное развитие благодаря тому, что его полный жизненный цикл протекает в теплое время года. По существующей классификации D. fasciola относится к морским, ведущим, сезонно-летним, олигосапробным макроводорослям Черного моря (Калугина-Гутник, 1975).
Выбор вида в качестве объекта исследования обусловлен его важностью как потенциального источника экономически важных полисахаридов. Повышенный интерес к этим соединениям объясняется наличием у них широкого спектра фармакологических свойств, низкой токсичностью для организма и возможностью получения на их основе лекарственных препаратов нового поколения (Меньшова, 2012). Экстрагирование D. fasciola показало наличие веществ, способных ингибировать рост морских бактерий и микроводорослей (Ktari et al., 2010).
При разработке рекомендаций для получения сырья из того или иного вида необходимо учитывать результаты популяционных исследований, которые дают наиболее полное представление о его роли в сообществе и устойчивости в нем. Особи, входящие в популяцию, в связи с различием в возрасте и жизненном состоянии отличаются друг от друга по мощности развития таллома и его отдельных частей, что так или иначе отражается на степени воздействия их на среду и другие организмы. Поэтому изучение размерно-массовой структуры ценопопуляций отдельных видов полезно для понимания особенностей формирования бентосных сообществ (Евстигнеева, Танковская, 2017в).
Известно, что популяции растений отличаются от популяций животных тем, что первичные адаптации у животных обеспечиваются сменой их поведения, а у растений они реализуются за счет изменений динамики роста, числа метамеров, типа структуры различных органов и др. (Злобин и др., 1986). Это подчеркивает необходимость применения специальных методов исследования популяций, в том числе морфометрического. Проводимые в данном ключе исследования в Черном море малочисленны и охватывают незначительное число видов (Евстигнеева, Танковская, 2017 а, б, в). В связи с этим целью работы стало изучение размерно-массовых характеристик слоевища и ценопопуляции D. fasciola, определение степени и характера их пространственно-временной изменчивости. Ранее подобные исследования черноморской D. fasciola не проводились.
Материал и методы
Основой данного исследования послужили альгологические сборы, произведенные в период с 2002 по 2016 гг. Отбор материала проводили вручную методом пробных площадок 25 х 25 см (Калугина, 1969) с последующим отбором 100 и более слоевищ. На рис. 1 представлена карта-схема районов выполнения работ.
Исследования проводили по двум направлениям: пространственное (один сезон – разные районы) и временное (а. один район – помесячно в течение вегетационного периода; б. один сезон – один район – разные годы). Для анализа были выбраны следующие параметры: длина (l), масса (m), ширина (sh) слоевища, количество размерно-массовых и модальных классов, частота встречаемости и количество (n) дихотомических разветвлений («вилки»), тип размерно-массового спектра (дискретный или непрерывный, моно- или мультимодальный), характер смещения доминирующих классов, мода (Мо), средние величины размерно-массовых показателей (хср.), лимиты (Lim) и размах (R) вариации, доверительный интервал (Δ), коэффициенты вариации (Cv) и корреляции (r) (Шмидт, 1985; Шитиков и др., 2003). Силу и характер изменчивости параметров определяли по пятибалльной шкале Г.Н. Зайцева (Зайцев, 1990).
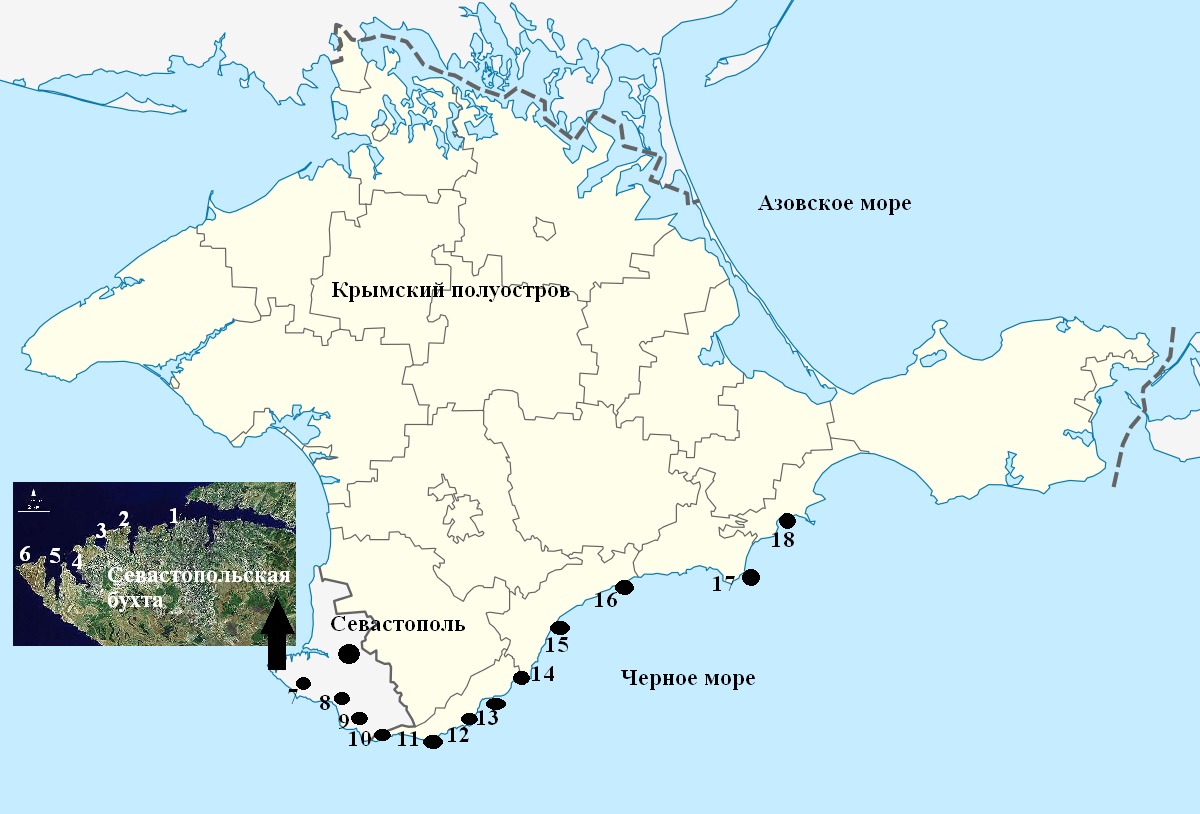
Рис. 1. Карта-схема районов исследования:
1 – бухта Мартынова, 2 – Парк Победы, 3 – бухта Круглая, 4 – бухта Камышовая, 5 – бухта Казачья, 6 – мыс Херсонес, 7 – мыс Фиолент, 8 – бухта Балаклавская (Золотой пляж), 9 – мыс Айя, 10 –урочище Батилиман, 11 – поселок Форос, 12 – поселок Кацивели, 13 – поселок Симеиз,
14 – мыс Мартьян, 15 – поселок Партенит, 16 – поселок Рыбачье, 17 – город Судак, 18 – Карадаг
Fig. 1. Scheme (map) of the region of investigation:
1 – the Martyn bay, 2 – the Victory park, 3 – the Kruglaya bay, 4 – the Kamyshovay bay, 5 – the Kazachya bay, 6 – the cape Chersonese, 7 – the cape Fiolent, 8 – the Balaklava bay, 9 – the cape Ajja , 10 – the tract Batiliman, 11 – the village Foros, 12 – the village Katsiveli, 13 – the village Simeiz, 14 –
the cape Martyan, 15 – the village Partenit, 16 – the village Fishing, 17 – the city Sudak,
18 – the Karadag
Многолетние и сезонные изменения параметров ценопопуляции и слоевища D. fasciola исследовали на примере поселений вида в урочище Батилиман, в бухте Балаклавская и у мыса Херсонес. Для оценки степени и направленности пространственных трансформаций этих параметров воспользовались альгологическими сборами в 18 районах черноморского побережья Крыма (см. рис. 1).
Размерно-массовую структуру ценопопуляции определяли путем взвешивания и измерения слоевищ во влажном состоянии. При группировании полученных данных число классов и классовый интервал выявляли в соответствии с формулой Стерджеса (Sturges, 1926). На основе этой формулы для вариационного ряда параметров D. fasciola приняли шаг 1,0 см для длины слоевища, 0,5 см для его ширины до первого разветвления, 0,05 г для массы. Число вариант, относящихся к тому или иному классу, выражали в процентах от объема выборки. Статистическую обработку материала проводили с использованием пакета программ STATISTICA 6.0. Различия считали достоверными на уровне значимости p ≤ 0,05.
Результаты и обсуждение
Пространственная динамика размерно-массовых параметров слоевища и ценопопуляции D. fasciola в летний и осенний периоды вегетации
Летний период. Размерные параметры. В ценопопуляции вида на исследованных участках Крымского прибрежья Черного моря индивидуальная длина слоевища варьирует в широких пределах (0,5–14,7 см). Размах пространственных изменений этого показателя составляет 2,6–13, 2 см с максимумом в прибрежье поселка Кацивели и в среднем достигает 5,6±1,1 см.
Средняя для каждого района длина слоевища максимальна в прибрежье Парка Победы (бухта Круглая) и минимальна – у мыса Херсонес. Оба района существенно отличаются по интенсивности циркуляции морских вод и ветрового волнения. Динамика окружающей морской среды требует от представителей литоральных экосистем адаптации своих биомеханических свойств к агрессивным внешним условиям (Herrel et al., 2006). При этом наиболее очевидными изменениями являются уменьшение общей площади и размеров объекта, укрепление его внешней оболочки. Подтверждением этого и является пространственная приуроченность крайних значений длины слоевища диктиоты. В каждом районе длина особей варьирует преимущественно по «нормальному» типу. Для всего массива полученных данных средняя длина слоевища составляет 3,9±0,3 см, а сам показатель отличается уровнем изменчивости, более высоким, чем в границах каждого района (Cv = 47%).
Количество классов, входящих в размерный спектр ценопопуляции D. fasciola, колеблется от 3 до 10 и в среднем достигает 6,0±0,9. Пространственная изменчивость этого параметра невелика и соответствует «верхней» норме. Максимально разнообразен размерный спектр в бухте Казачья и вблизи поселка Симеиз. Близок к нему и состав ценопопуляции в акватории города Судак. Ценопопуляция, независимо от района обитания, обладает такими одинаковыми качествами размерного спектра, как его преимущественная мономодальность, непрерывность и поливариантность размещения модального класса (чаще право- и левостороннее, реже – его отсутствие).
Как правило, среди размерных классов преобладает какой-то один (модальный класс). На его долю приходится 24–56% общего числа особей. Среди значений моды чаще встречаются 3 и 5 см. Особи более крупных размеров (9 см и более) зарегистрированы в районе поселка Кацивели и бухты Казачья, однако их вклад в общий состав невелик.
Существенным разбросом характеризуются значения ширины слоевища (0,05–0,5 см). Особенно велико различие ее крайних величин у особей вида в бухте Камышовая. Средний для всего обследованного берега размах колебаний ширины слоевища достигает 0,2±0,04 см, а его изменчивость в пространстве находится в пределах «верхней» нормы (Cv = 40%). Ширина, средняя для особей каждого района, характеризуется большей стабильностью, чем их длина. Ее максимум приурочен сразу к нескольким районам, что, впрочем, присуще и ее минимуму. Изменчивость показателя в каждом районе и по всему массиву данных по шкале Г.Н. Зайцева является «верхненормальной».
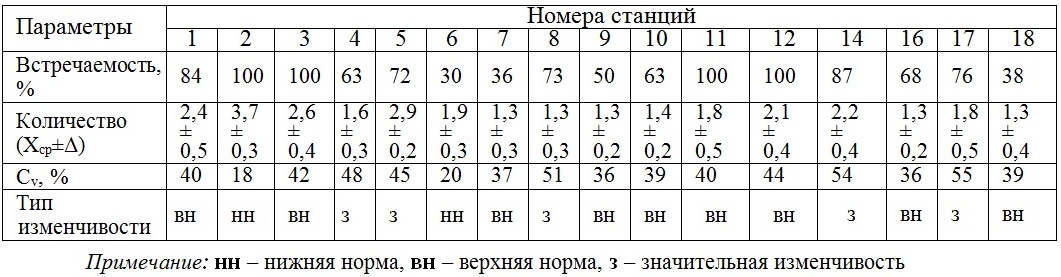
Дихотомические разветвления слоевища. Встречаемость дихотомических разветвлений («вилок») на слоевищах в разных районах неодинакова: 30–100%. Среднее число «вилок» у их обладателей изменяется от 1,3 до 3,7. Вариабельность параметра в каждом районе соответствует «норме» и изредка – «значительному» типу, который, в свою очередь, характерен и для изменений анализируемого параметра вдоль всех берегов (табл. 1).
Таблица 1. Встречаемость и показатели пространственной изменчивости числа дихотомических разветвлений у D. fasciola
Table 1. Occurrence and indexes of spatial variability number of D. fasciola dichotomizing branching
Массовые параметры. Масса слоевища D. fasciola вдоль исследованных берегов изменяется очень широко (от 0,001 до 0,8 г). Наибольшее различие крайних значений параметра отмечено в районе мыса Фиолент. Близок к нему и такой же размах величин массы у особей в бухте Круглая. В районе поселков Партенит, Рыбачье, Симеиз, у Карадага, в бухтах Балаклавская и Мартынова различие крайних значений массы слоевища несущественно.
Средняя масса слоевища особей в каждом районе варьирует от 0,01±0,001 до 0,13±0,04 г. Для всего массива данных этот показатель составляет 0,04±0,004 г. Его изменчивость более интенсивная, чем у длины. В большинстве районов она проявляется как «значительная», «большая» и «очень большая». Высокая изменчивость массовых параметров слоевища и ценопопуляции вида является отражением закономерности, характерной для других биологических объектов, описанной И.И. Шмальгаузеном (Шмальгаузен, 1935).
Количество классов, входящих в массовый спектр ценопопуляции, составляет 1–5. Самое высокое разнообразие спектра обнаружено на тех участках, где велики и средние значения массы слоевища (Парк Победы и бухта Круглая). Количество массовых классов вдоль исследованного берега колеблется в «значительной» степени, превышающей таковую у размерных классов. Почти в половине районов массовый спектр состоит из особей единственного класса с массой 0,05 г. Встречаемость такой моды очень высокая. Кроме нее возможны и такие варианты, как 0,1 и 0,2 г. Вклад растений лидирующих классов достигает 19–100%. Минимум доли особей модального класса приходится на ценопопуляцию с высоким разнообразием массового состава. Структурообразующая роль крупновесовых особей незначительна.
В случаях, когда спектр включает более одного класса, возможны следующие варианты размещения модального класса и полночленности массового спектра: преимущественно левоассиметричное, эпизодически правоассиметричное, равный вклад непрерывного и дискретного типов спектра.
В целом, статистическая закономерность модификационной изменчивости, соблюдаемая в отношении распределения значений длины и ширины слоевища, нарушается в случае массовых параметров за счет высокого вклада особей наименьшего класса.
Анализ данных, представленных в табл. 2, позволяет оценить силу и направленность взаимосвязи изученных признаков D. fasciola.
Таблица 2. Коэффициенты корреляции взаимозависимых изменений некоторых размерно-массовых параметров D. fasciola в разных районах Крымского прибрежья
Table 2. Correlation coefficients of interdependent changes of some dimensional and mass D. fasciola parameters in different areas Crimea coast
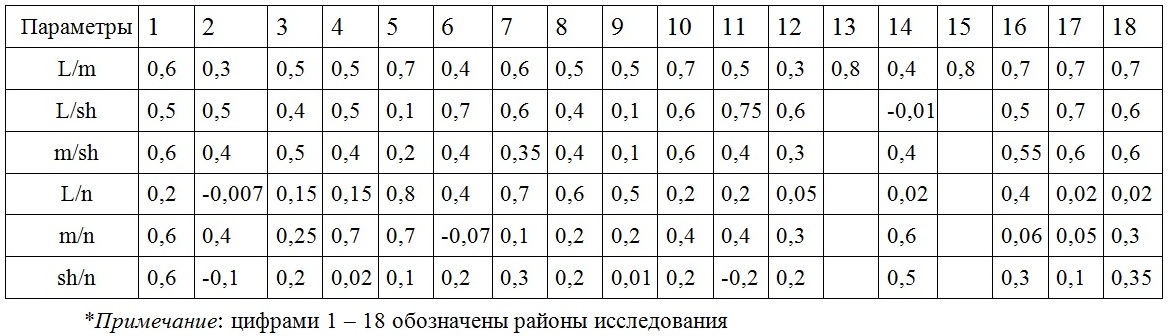
Летом для сопряженных изменений длины и массы слоевища на всем протяжении исследованных берегов характерна положительная корреляция, среди типов которой преобладают слабая и средняя связь (r = 0,3–0,7). Максимум выражения такой связи проявляется у особей в акватории поселков Партенит и Кацивели. Примерно такой же является корреляционная связь взаимных изменений длины и ширины, массы и ширины слоевища. Очень слабой выглядит сопряженная связь ширины и количества «вилок».
Осенний период. D. fasciola относится к группе сезонно-летних водорослей Черного моря. Наши исследования показали, что этот вид является существенным компонентом фитобентоса и в осенний период. Следовательно, появляется смысл определить размерно-массовые параметры ценопопуляции и слоевища, их внутрипопуляционную изменчивость в указанный период. В последующем возможно сравнение данных за два сезона в одних и тех же районах (табл. 3).
Таблица 3. Сравнительная характеристика размерно-массовых параметров D. fasciola летом и осенью в разных районах
Table 3. The comparative characteristic of the D. fasciola dimensional and mass parameters in the summer and fall in different areas
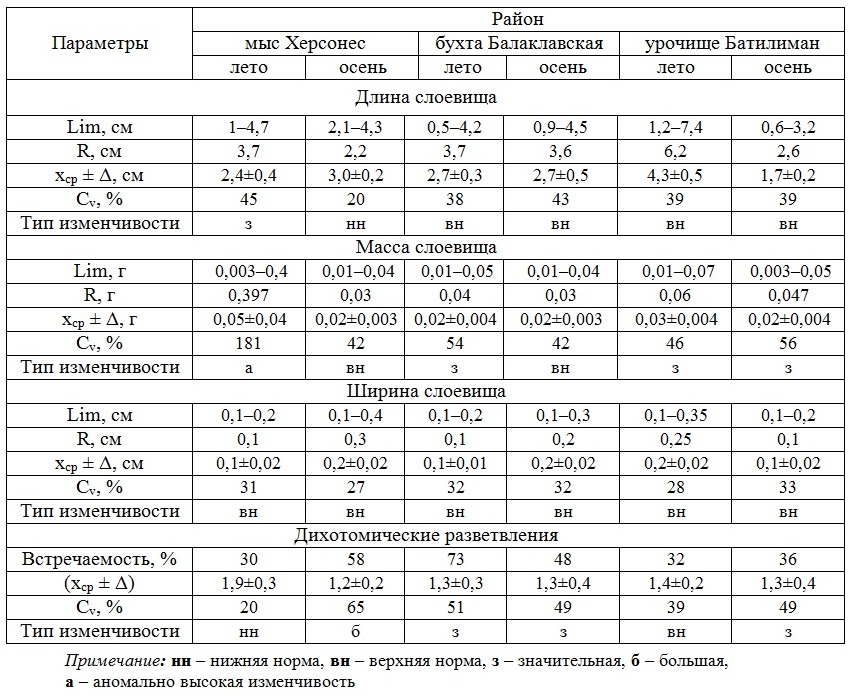
В ходе исследования было установлено, что в прибрежье мыса Херсонес, бухты Балаклавская и урочища Батилиман наибольшее межсезонное сходство анализируемых параметров проявлялось у особей вида в бухте и наименьшее – в районе мыса. Обе акватории существенно отличаются степенью защищенности берегов. Для поселений диктиоты в урочище Батилиман и в районе мыса Херсонес общим являются непрерывность размерного спектра и левоассиметричность спектра значений ширины слоевища. В бухте Балаклавская и в урочище Батилиман длина растений варьировала по одному и тому же «верхненормальному» типу. Для особей ценопопуляции трех районов в оба сезона был характерен одинаковый тип изменчивости ширины их слоевища.
Вместе с тем, полученные данные свидетельствуют об отсутствии какого-либо межсезонного сходства по таким параметрам ценопопуляции в каждом районе, как тип смещения модальных классов и степень дискретности массового состава. Более того, в бухте Балаклавская и у мыса Херсонес в разные сезоны нет аналогии в дислокации сгущения значений ширины слоевища.
Среди трех районов в бухте Балаклавская ценопопуляция летом и осенью проявляет больше сходства, чем отличий. К последним следует отнести более высокие в летний период изменчивость количества «вилок» и их встречаемость, а осенью – размах вариабельности и среднее значение ширины слоевища, количество классов в спектре ее величин. По остальным показателям наблюдалось абсолютное или близкое к нему совпадение. В двух других районах различие параметров D. fasciola в оба сезона является более существенным. Например, летом в акватории урочища Батилиман уровень очень многих параметров превышает таковой осенью. Для осеннего периода была характерна лишь повышенная изменчивость количества «вилок». Летние максимумы многих размерно-массовых параметров в прибрежье урочища Батилиман и мыса Херсонес совпадают.
В целом, в урочище Батилиман летний уровень большинства анализируемых характеристик превосходит осенний, в бухте Балаклавская ситуация имеет обратный характер, а в прибрежье мыса Херсонес для ценопопуляции в оба сезона характерна примерная паритетность проявления максимумов.
Усреднение сезонных данных для каждого из трех районов показало, что длина и масса слоевища, размах их крайних значений, разнообразие спектров, среднее количество «вилок» выше летом. Ширина слоевища осенью немного больше (на 30%), зато размах ее значений, количество классов в соответствующем спектре и встречаемость дихотомических разветвлений в оба сезона практически совпадают. Эти результаты свидетельствуют о том, что несмотря на более продолжительный, чем принято считать, срок вегетации, лето остается для вида сезоном максимумов. Осенью наблюдается скорее рост в ширину, чем в длину. Внутрипопуляционная изменчивость значений длины, количества размерных классов и разнообразие классов значений ширины слоевища осенью и летом, как правило, одинаковая. Изменчивость массы слоевища выше летом, в период массовой вегетации вида.
Внутригодовая изменчивость размерно-массовых параметров слоевища и ценопопуляции D. fasciola
В табл. 4 представлены данные, свидетельствующие о том, что максимальный уровень параметров слоевища и ценопопуляции приходится на вторую половину года и, прежде всего, на июль и август. В динамике длины слоевища, размаха ее ежемесячных вариаций в период с января по сентябрь наблюдается постепенное увеличение показателей, а затем их спад. Ежемесячно, за небольшим исключением, длина слоевища меняется в пределах «верхней» нормы, а сам спектр размерных классов является непрерывным. Величина моды в конце осени и зимой невелика (2 и 3 см), к этим же периодам вегетации приурочено высокое долевое участие особей модального класса, а значит и более выраженная однородность размерного состава (рис. 2). Спектр размерных классов отличается как симметричностью в расположении доминирующих классов, так и преимущественным смещением их влево.
Таблица 4. Изменчивость размерно-массовых параметров D. fasciola по месяцам
Table 4. Variability of the D. fasciola dimensional and mass parameters on months
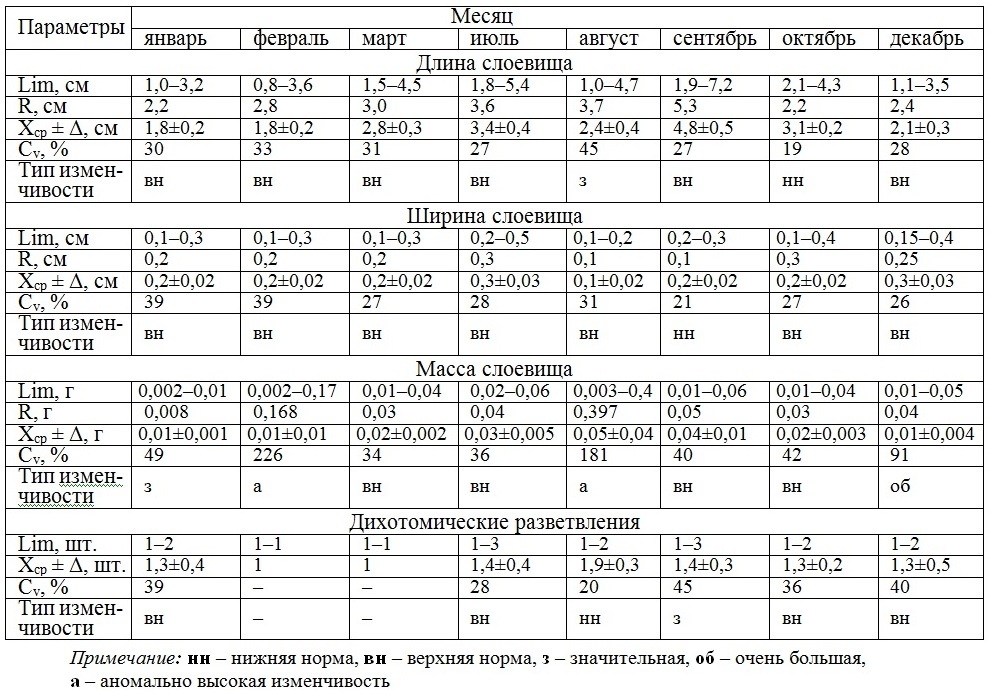
Ширина слоевища варьирует по месяцам в пределах «верхней» нормы. Ее среднее значение в разные периоды отличается мало. Более существенными выглядят вариации числа классов, входящих в соответствующий спектр величин ширины слоевища, который, к тому же, может быть как непрерывным, так и дискретным, симметричным или очень близким к такому типу.
В динамике массы слоевища отмечено возрастание ее с января по август, после чего происходит снижение, особенно в октябре и декабре. Более резко меняется размах крайних значений массы. Ежемесячные вариации индивидуальной массы растений происходят в соответствии со шкалой изменчивости биологических признаков по 4 типам, среди которых преобладают более умеренные «верхненормальный» и «значительный» (Cv = 34–49%). Количество массовых классов изменяется от 1 до 5, но чаще спектр состоит из одного или двух классов (рис. 2). Доля модальных массовых классов существенно выше размерных. В отличие от последнего мода в массовом спектре одна и та же. Все это указывает на высокую однородность массового состава ценопопуляции D. fasciola. Массовый спектр всегда левоассиметричный, сочетающий в себе как непрерывность, так и дискретность распределения особей по классам.
В пределах «верхней» нормы ежемесячно меняется количество разветвлений, средняя величина которого большую часть вегетационного периода остается одинаковой и только в августе становится в 1,5 раза выше.
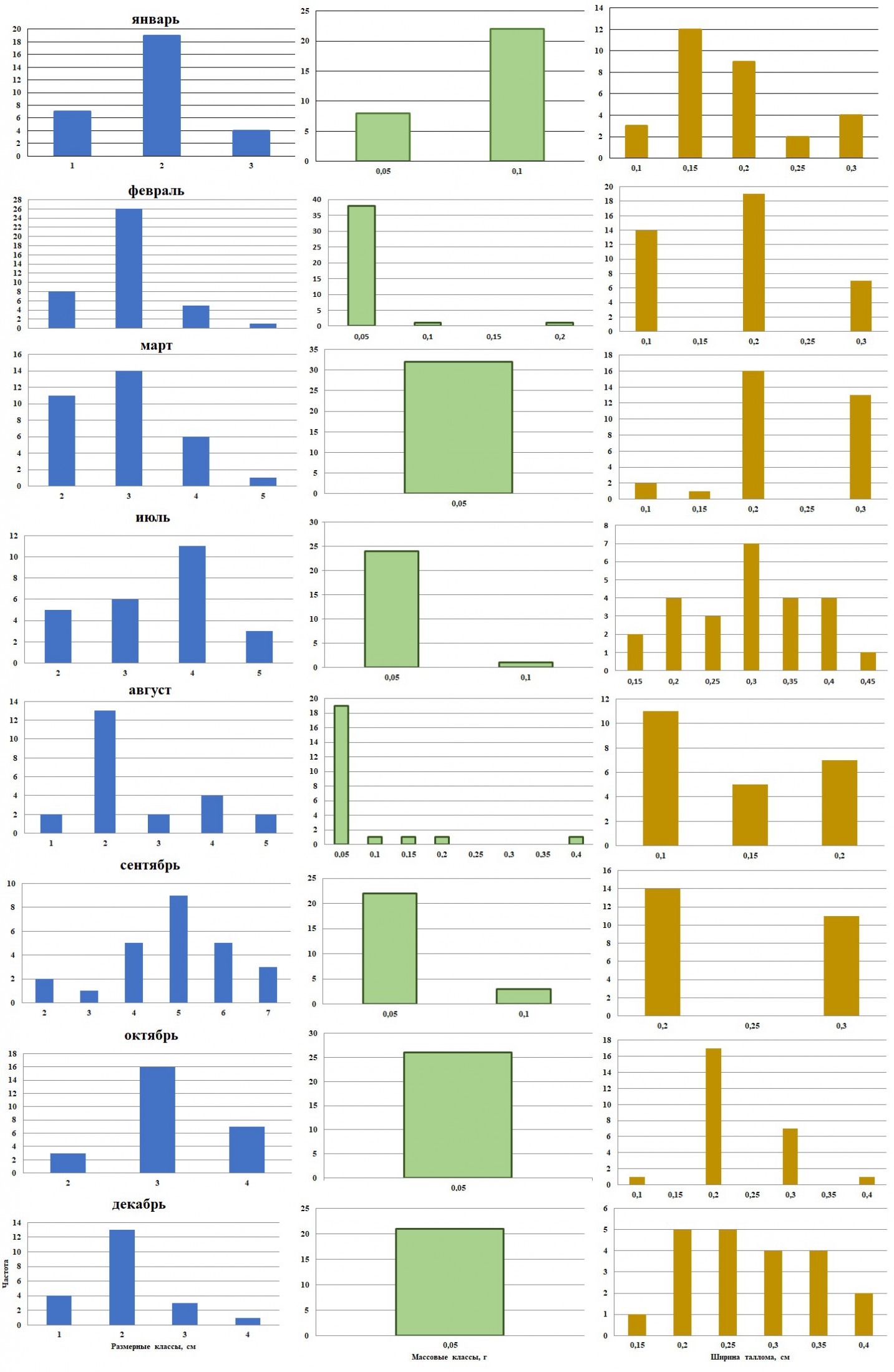
Рис. 2. Внутригодовая изменчивость частоты встречаемости размерно-массовых классов в ценопопуляции D. fasciola
Fig. 2. Intra annual variability of occurrence frequency dimensional and mass classes in D. fasciola cоenopopulation
Разногодичная изменчивость размерно-массовых параметров слоевища и ценопопуляции D. fasciola
В табл. 5 сведены данные, свидетельствующие об особенностях разногодичной изменчивости слоевища и ценопопуляции D. fasciola в одном и том же районе. Установлено, что значения анализируемых параметров наиболее высоки в первый год фиксированных наблюдений и низки – в последний.
Таблица 5. Разногодичная изменчивость размерно-массовых параметров D. fasciola
Table 5. Interannual variability of the D. fasciola dimensional and mass parameters
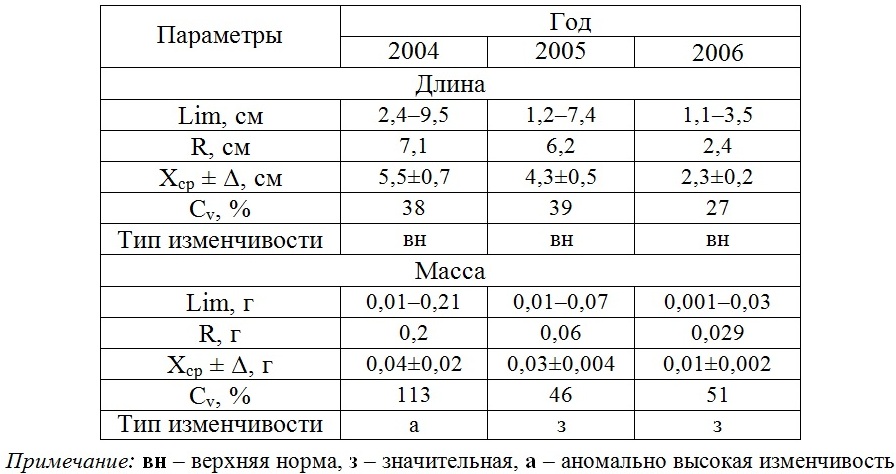
Различие крайних значений длины и массы слоевища достигает 2 и 4 раз соответственно. При этом внутрипопуляционная вариабельность длины слоевища в разные годы остается одинаковой, у массы же в начале наблюдений она высокая и менее интенсивная в последующем. Умеренной является и межгодовая изменчивость числа классов в каждом спектре. Оба спектра полночленные, с разными вариантами сгущения доминирующих классов. Доля модального размерного класса существенно ниже массового (рис. 3).
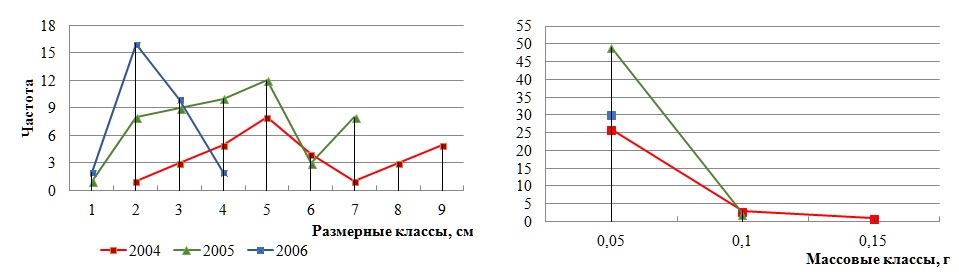
Рис. 3. Разногодичная изменчивость частоты встречаемости размерно-массовых классов в ценопопуляции D. fasciola (урочище Батилиман)
Fig. 3. Interannual variability of occurrence frequency of dimensional and mass classes in D. fasciola coenopopulation (the tract Batiliman)
Таким образом, отмечено снижение длины и массы слоевища, а также разнообразия классов в спектрах от начала (2004 г.) до конца фиксированных наблюдений (2006 г.). Для обоих спектров наиболее характерны непрерывность и левоассиметричность. Изменчивость массы неизменно выше, чем длины слоевища. Вклад модального массового класса в общую структуру в несколько раз больше размерного, что указывает на однородность массового состава.
В целом, у D. fasciola изменения длины слоевища осуществляются по двум типам с преобладанием «верхненормального» подтипа, вклад которого составляет 61–100% с максимумом у разногодичной вариабельности. Пространственные и разногодичные изменения массы D. fasciola превышают «норму». Особенностями внутригодовой изменчивости массы слоевища является преобладание ее «верхненормального» подтипа, доля которого достигает 50%, однако он компенсирован вкладом изменчивости, превышающей «норму» для биологических признаков. Размерные спектры всегда или в большинстве случаев являются непрерывными, массовые – непрерывными в течение всего периода вегетации и в разные годы, но у ценопопуляции в отдельных районах они могут быть как непрерывными, так и дискретными. Размерно-массовые спектры D. fasciola преимущественно левоассиметричные.
Выводы
Выявлена пространственная локация максимумов и минимумов параметров слоевища и разнообразия размерно-массовых спектров ценопопуляции D. fasciola. Показано, что среди количественных признаков наименее подвержены изменениям в пространстве размерные характеристики, наиболее – массовые.
Установлено, что в процессе вегетации с января по сентябрь происходит плавное увеличение растений в длину, после чего этот процесс приостанавливается. Средние значения ширины слоевища и количества дихотомических разветвлений большую часть вегетационного периода остаются неизменными. Внутригодовая динамика массы и длины слоевища в разные месяцы во многом тождественна.
Особенностями сезонной изменчивости параметров слоевища являются летний максимум его длины, массы, количества дихотомических разветвлений и разнообразия размерно-массовых спектров ценопопуляций. Для осеннего периода характерен интенсивный рост слоевища в ширину.
Разногодичная динамика параметров D. fasciola проявляется в снижении длины, массы слоевища и разнообразия соответствующих классов в ценопопуляции с первого по последний год фиксированного периода наблюдений (2004–2006 гг.).
Показано, что независимо от места обитания и времени вегетирования размерные спектры D. fasciola всегда или в большинстве случаев непрерывные, массовые – как непрерывные, так и дискретные, но преимущественно левоассиметричные.
Работа выполнена по теме госзадания ФГБУН ИМБИ РАН «Исследование механизмов управления продукционными процессами в биотехнологических комплексах с целью разработки научных основ получения биологически активных веществ и технических продуктов морского генезиса» номер госрегистрации АААА-А18-118021350003-6.
Список литературы
- Евстигнеева И.К., Танковская И.Н. Анализ природных ценопопуляций Gelidium spinosum (S.G. Gmelin) P.C. Silva (Rhodophyta) в прибрежье Черного моря // Водные биологические ресурсы России: состояние, мониторинг, управление: материалы Всероссийской научн. конф. с межд. участием, посвященной 85-летию Камчатского научно-исследовательского института рыбного хозяйства и океанографии (3–6 октября 2017 г., Петропавловск-Камчатский). – КамчатНИРО. 2017 а. – С. 214–222. URL: http://www.kamniro.ru/files/2017.pdf
- Евстигнеева И.К., Танковская И.Н. Размерно-массовые характеристики слоевища и ценопопуляций Ulva linza L. (Chlorophyta) и их динамика в Черном море // Вопросы современной альгологии. 2017 б. №2 (14) URL: http://algology.ru/1175 (дата обращения – 1.10.2018)
- Евстигнеева И.К., Танковская И.Н. Размерно-массовые характеристики слоевища и ценопопуляций Ulva intestinalis L. (Chlorophyta) и их динамика в Черном море // Вопросы современной альгологии. 2017 в. №2 (14). URL: http://algology.ru/1174 (дата обращения – 1.10.2018)
- Зайцев Г.Н. Математика в экспериментальной ботанике. – М.: Наука, 1990. – 296 с.
- Зинова А.Д. Определитель зеленых, бурых и красных водорослей южных морей СССР. –М., Л.: Наука, 1967. – 397 с.
- Злобин Ю.А., Кохановский В.М., Сухой И.Б. Ценопопуляционный уровень в решении проблем фитоценологии // Флора и растительность Украины. – Киев, 1986. – С. 41–43.
- Калугина А.А. Исследование донной растительности Черного моря с применением легководолазной техники // Морские подводные исследования. – М., 1969. – С. 105–113.
- Калугина-Гутник А.А. Фитобентос Черного моря. – Киев: Наук. думка, 1975.– 248 с.
- Меньшова Р.В. Полисахариды некоторых видов бурых водорослей: Автореф. дис. канд. хим. наук. – Владивосток, 2013. – 23 с.
- Танковская И.Н., Евстигнеева И.К. Морфоструктура бурой водоросли Dilophus fasciola Roth Howe в прибрежных фитоценозах района Севастопольской бухты // Экология моря. 2002. Вып.60. С. 12–15.
- Шитиков В.К., Розенберг Г.С., Зинченко Т.Д. Количественная гидроэкология: методы системной идентификации. – Тольятти: ИЭВБ РАН, 2003. – 463 с.
- Шмальгаузен И.И. Определение основных понятий и методика исследования роста. Рост животных. – М.: Биомедгиз, 1935. – С. 8–60.
- Шмидт В.М. Развитие представлений о корреляциях и корреляционной структуре биологических объектов // Исследование биологических систем математическими методами. Труды Биол. НИИ ЛГУ. 1985. №37. С. 5–18.
- Guiry M.D., Guiry G.M. AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org (дата обращения – 16.10.2018).
- Herrel A., Speck T., Rowe N. Ecology and Biomechanics: A Mechanical Approach to the Ecology of Animals and Plants. – Boca Raton, CRC Taylor & Francis. Available from: 2006. – 334 p.
- Ktari L., Ismail-Ben Ali A., Ben Redjem Y., Langar H., El Bour M. Antifouling activity and chemical investigation of the brown alga Dictyota fasciola (Dictyotales) from Tunisian coast // Cah Biol Mar. 2010. 51. P. 109–115.
- Sturges H. The choice of a class-interval // Journal American Statistical Association. 1926. Т.21. №153. P. 65–66.
Статья поступила в редакцию 25.10.2018
Dimension-mass properties of the thallus and coenopopulations
of the Dictyota fasciola (Roth) J.V. Lamouroux (Ochrophyta)
and their variability in the Black Sea
Irina K. Evstigneeva, Irina N.Tankovskaya
Kovalevsky Institute of Marine Biological Research of RAS (Sevastopol, Russia)
As a result of long-term studies the size-mass parameters of thallus and cоenopopulations of the Black Sea macroalgae Dictyota fasciola (Roth) J.V. Lamouroux (Ochrophyta) were determined for the first time. Mean and extreme values, as well as spatial-temporal localization and variability of these parameters were estimated. Constant and dynamical signatures of thallus and coenopopulations were established.
Key words: Dictyota fasciola (Roth) J.V. Lamouroux; Ochrophyta; thallus; coenopopulation; dimension-mass structure; dynamics; Crimea; Black Sea.
Об авторах
Евстигнеева Ирина Константиновна - Evstigneeva Irina K.
кандидат биологических наук
старший научный сотрудник, Институт морских биологических исследований имени А.О.Ковалевского РАН, Севастополь, Россия (Kovalevsky Institute of Marine Biological Research of RAS, Russia, Sevastopol)
ikevstigneeva@gmail.com
Танковская Ирина Николаевна - Tankovskaya Irina N.
младший научный сотрудник, Институт морских биологических исследований имени А.О.Ковалевского РАН, Севастополь, Россия (Kovalevsky Institute of Marine Biological Research of RAS, Russia, Sevastopol)
logrianin@nm.ru
Корреспондентский адрес: Россия, 299011, Севастополь, пр. Нахимова, 2, ФГБУН ИМБИ; тел. 88692550795.
ССЫЛКА НА СТАТЬЮ:
Евстигнеева И.К., Танковская И.Н. Размерно-массовые характеристики слоевища и ценопопуляции Dictyota fasciola (Roth) J.V. Lamouroux (Ochrophyta) и их изменчивость в Черном море // Вопросы современной альгологии. 2018. № 3 (18). URL: http://algology.ru/1327
Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно.
Адрес - info@algology.ru
При перепечатке ссылка на сайт обязательна
На ГЛАВНУЮ
Карта сайта
К разделу ОБЗОРЫ, СТАТЬИ И КРАТКИЕ СООБЩЕНИЯ

|
