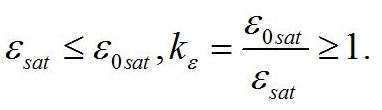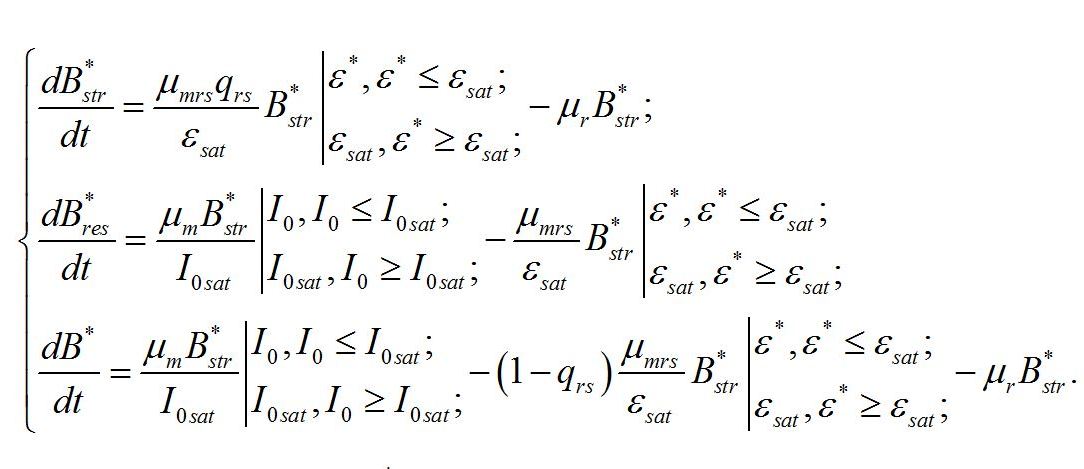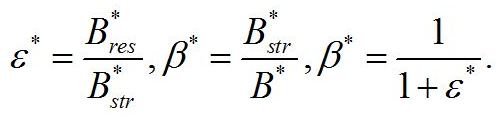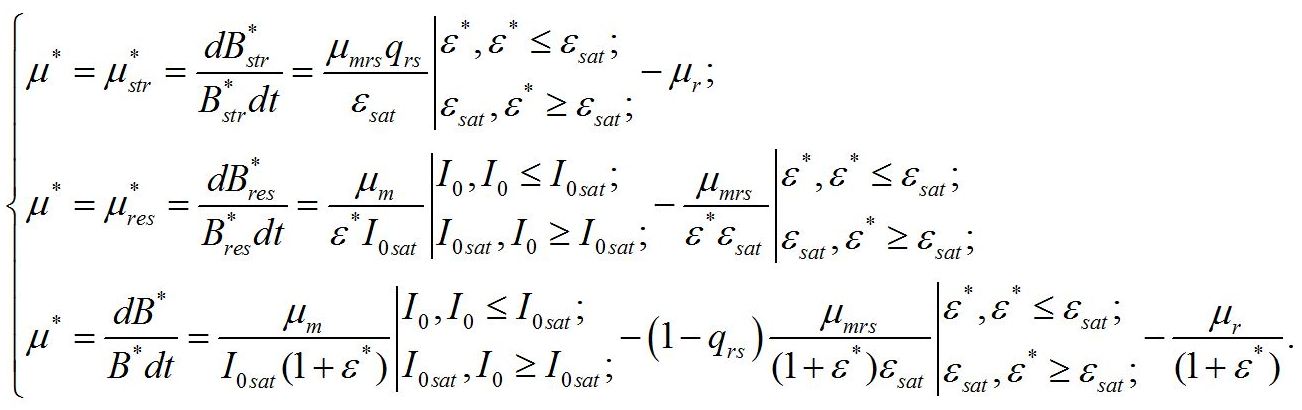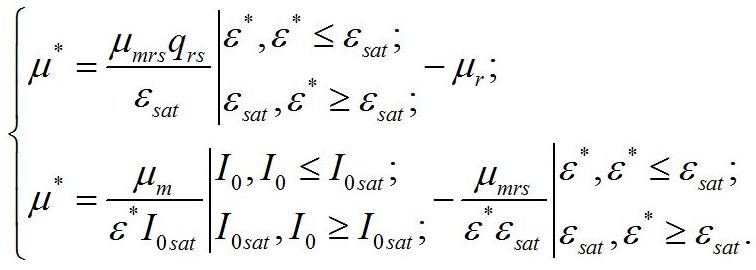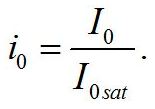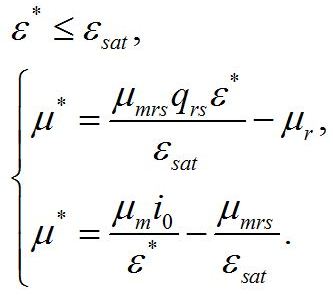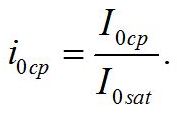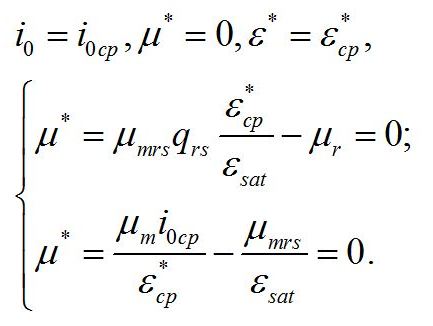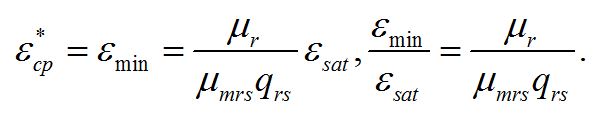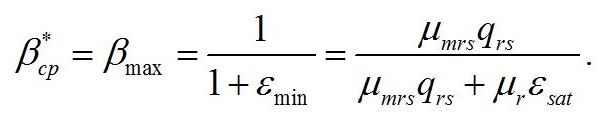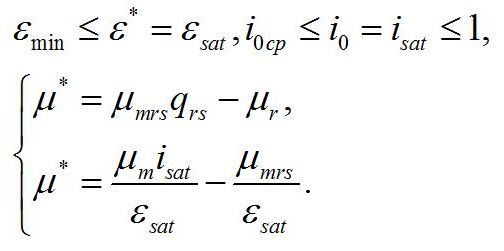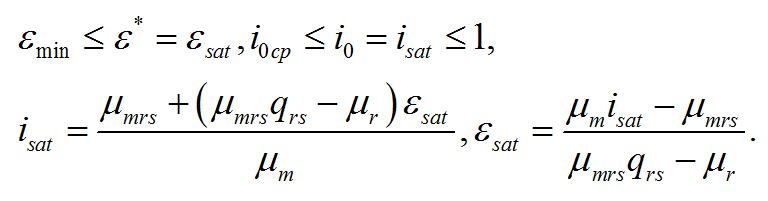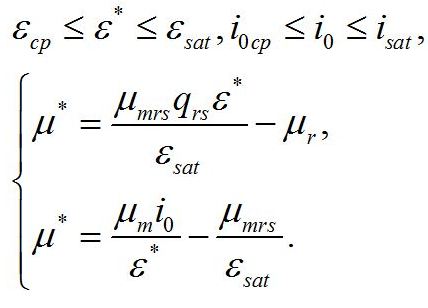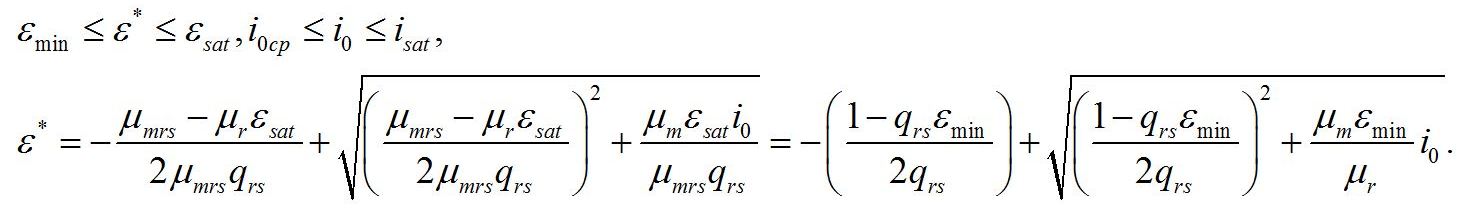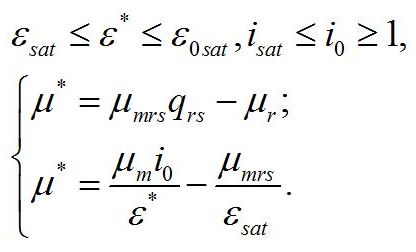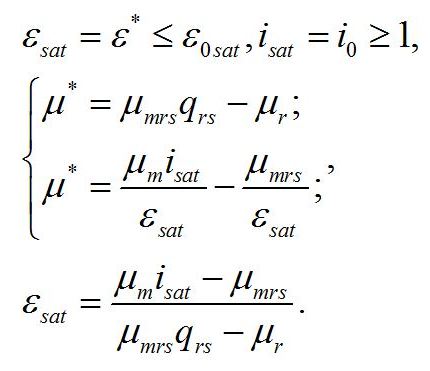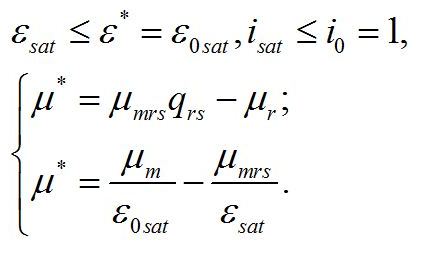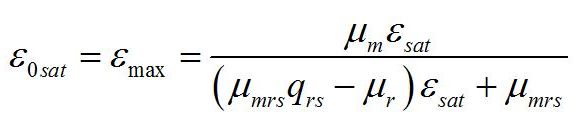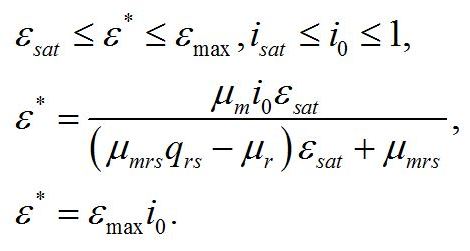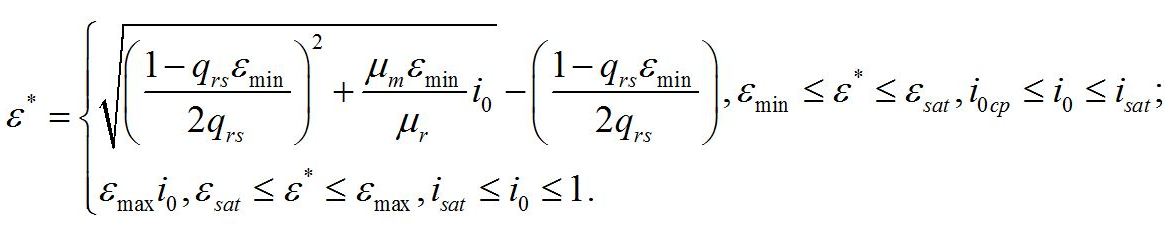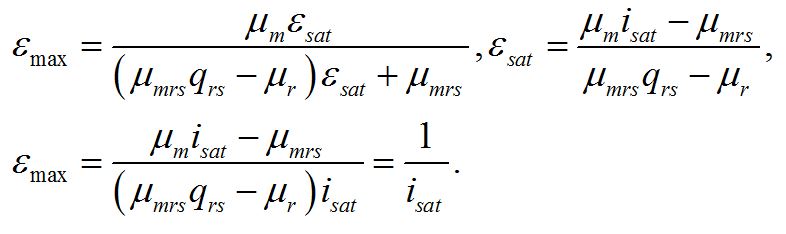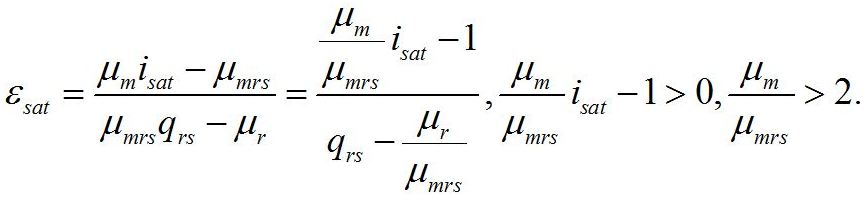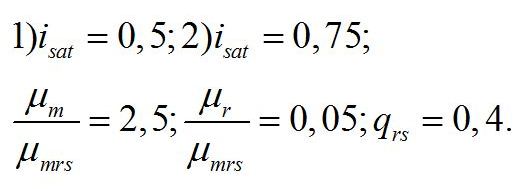|
|
|
Влияние света на макромолекулярный состав микроводорослей в непрерывной культуре невысокой плотности (Часть 2)
Тренкеншу Р.П. Rudolf P. Trenkenshu
Институт морских биологических исследований им. А.О. Ковалевского (г. Севастополь)
УДК 582.232:581.121
Показано, что форма зависимости макромолекулярного состава биомассы микроводорослей от света определяется соотношением насыщающих значений фотосинтеза и синтеза структурных компонентов клеток. При ограничении скорости процессов в последовательности преобразования резервных веществ в структурные, если при этом не достигнуто световое насыщение, в клетках накапливаются запасные вещества в виде углеводов и, возможно, липидов. Получены уравнения, позволяющие описать зависимость соотношения ресурсных (резервных) и структурных форм биомассы от интенсивности света в динамически равновесной (непрерывной) культуре невысокой плотности. Ключевые слова: микроводоросли; культивирование; скорость фотосинтеза; удельная скорость роста; биохимический состав.
Введение В первой части настоящей работы (Тренкеншу, 2017) рассматривалось влияние света на рост микроводорослей в культуре, как единственного внешнего лимитирующего фактора. Удалось установить, что при полном минеральном обеспечении клеток свет влияет на удельную скорость роста культуры опосредованно, через изменение соотношения ресурсных и структурных макромолекулярных форм биомассы клеток. Показано, что соотношение этих форм зависит как от внешнего света, так и от внутренних процессов в клетке. Эта опосредованность приводит к общепринятому разграничению понятия лимита скорости на лимитирующий фактор и узкое место или лимитирующее звено. Под лимитирующим фактором понимается поток (или концентрация) субстрата, который определяет скорость производства конечного продукта в цепи метаболических реакций до определенного предела (насыщение). Увеличение потока лимитирующего субстрата выше этого предела не приводит к увеличению скорости образования конечного продукта, т.е. скорость ограничена лимитирующим звеном. Такой подход согласуется с концепциями Либиха (Liebig, 1847), Блекмана (Blackman, 1905) и Друпа (Droop, 1973, 1979). Специфика влияния света на скорость фотоавтотрофного роста микроводорослей состоит в том, что фотоавтотрофный рост можно рассматривать, как минимум, в виде двухэтапного процесса (Физиология растений, 2005). На возможность такого деления особенно указывают работы, связанные с фотосинтезом листьев у высших растений (Ивлев, 2011). Первый этап представляет собой собственно фотосинтез, в результате которого в цикле Кальвина образуются углеводы. Второй этап представляет собой синтез структурных компонентов клетки из этих углеводов, выступающих в качестве источника энергии и углеродных скелетов. Это означает, что на разных этапах скорости регулируются различными лимитирующими факторами и звеньями. На первом этапе лимитирующим звеном является цикл Кальвина, на втором – один из ферментов или компонент транспортной системы, участвующий в последовательности преобразования углеродных скелетов в структурные компоненты клеток микроводорослей. В зависимости от того, какое звено является лимитирующим, свет будет по-разному влиять как на скорость роста микроводорослей, так и на макромолекулярный состав биомассы клеток. Как и ранее, предполагается, что все внешние факторы, кроме света, находятся на уровне, не влияющем на скорость роста. Внутриклеточное ограничение скорости роста. В первой части работы был рассмотрен случай лимитирования скорости роста микроводорослей светом, а лимитирующим звеном – цикл Кальвина. В данной работе анализируется случай, когда скорость ограничена только внутриклеточным лимитирующим звеном в последовательности преобразования углеводов в структурные компоненты клеток. Это звено характеризуется видоспецифическим параметром – максимальной величиной удельной скорости преобразования резервной биомассы в структурные формы. Какая ферментная или транспортная система ограничивает скорость процесса преобразования макромолекулярных форм биомассы у микроводорослей точно неизвестно. Укажем только, что для выяснения конкретного узкого места разработаны подходы, которые помогут выявить его локализацию (Рубин 2013; Минкевич, 2016). Для решения поставленной в данной работе задачи достаточно оперировать названным кинетическим видоспецифическим параметром. Принципиальным отличием второй части работы от первой является то, что рассматривается случай, когда лимитирующим звеном является не цикл Кальвина, а максимальная скорость трансформации резервных веществ в структурные формы. Это означает, что в таком случае удельная скорость роста микроводорослей достигнет насыщения при некоторой ненасыщающей интенсивности света, а соотношение резервные/структурные формы в клетках будет изменяться с ростом интенсивности света вплоть до светового насыщения. В данном случае:
Получение зависимости макромолекулярного состава микроводорослей от интенсивности света для такого случая является целью предлагаемой работы. Система уравнений. В основу работы положена система уравнений, предложенная ранее (Тренкеншу, 2016; Тренкеншу, 2017):
Надстрочные символы (*) означают, что рассматривается непрерывная динамически равновесная культура микроводорослей. В уравнениях приняты следующие обозначения. Биомасса микроводорослей представлена как сумма структурных и резервных (ресурсных) форм:
Соотношение этих форм:
Кинетические коэффициенты: - μrs – удельная скорость трансформации ресурсных веществ в структурные, - qrs – экономический коэффициент этого перехода, - μr – удельная скорость естественного распада структур, Учитывая, что в непрерывной культуре все удельные скорости равны и равны удельной скорости роста (или протока), получим:
В настоящей работе скорость роста микроводорослей, так же как и в первой части, отдельно не рассматривается. А величина удельной скорости роста используется только в качестве инструмента для решения систем уравнений. Учитывая, что третье уравнение является следствием первых двух, систему уравнений можно упростить:
Зависимость соотношения макромолекулярных форм биомассы от интенсивности света Система уравнений для ненасыщающих величин соотношений ε*. Введем для удобства безразмерную величину интенсивности света:
Для данных условий система редуцируется до вида:
Определим границы применимости для полученной системы уравнений. При отсутствии света применение уравнений не имеет смысла, т.к. в темноте невозможно получение непрерывной фотоавтотрофной культуры. Такой случай подробно рассмотрен в работе (Тренкеншу, 2016). Рост микроводорослей в фотоавтотрофной культуре возможен только при интенсивности света выше определенной величины, называемой компенсационным пунктом фотосинтеза (I0cp):
Световой компенсационный пункт. Этой точке на световой кривой роста будут соответствовать минимальные значения соотношения ресурсные/структурные формы биомассы. Скорость роста будет равной нулю:
Из уравнений находим величину соотношения ресурсные/структурные формы биомассы в точке, соответствующей световому компенсационному пункту при фотоавтотрофном росте микроводорослей. В этом случае величина соотношения будет минимальной для непрерывной культуры. Из первого уравнения системы находим соотношение ресурсные/структурные формы биомассы в световой компенсационной точке:
Содержание структурных форм органического вещества в биомассе клеток в световой компенсационной точке будет максимальным. Вводя соответствующие обозначения, получим:
Второе уравнение позволяет выразить величину светового компенсационного пункта через кинетические коэффициенты:
Найденная величина интенсивности света является нижней границей применения системы уравнений. Верхней границей служит интенсивность света (Isat), при которой наступает ресурсное насыщение, т.е. лимитирующим звеном служит ферментная или транспортная система в цепи превращения резервных веществ в структурные (εsat):
При достижении ресурсного насыщения система уравнений запишется в виде:
Решение системы позволяет найти интенсивность света (isat), при которой наступает ресурсное насыщение, или выразить ресурсное насыщение через величину интенсивности света, при которой наступает это насыщение:
Эти величины представляют собой верхнюю границу применимости системы уравнений:
Зависимость соотношения ресурсных и структурных макромолекулярных форм в биомассе микроводорослей от интенсивности света в пределах найденных границ будет описываться таким же уравнением, как и в первой части работы:
Система уравнений для интенсивностей света в диапазоне isat £ i0 £ 1. При интенсивности света выше найденной границы isat, соотношение ресурсные/структурные формы будет зависеть от интенсивности света, вплоть до насыщающей интенсивности i0 = 1, при которой установится соотношение ε0sat. Для этого диапазона освещенностей система уравнений имеет вид:
Решение системы в точке смены лимитирующих звеньев (εsat) является нижней границей возможности использования данной системы уравнений:
Как и следовало ожидать, последняя формула совпадает с верхней границей предыдущей системы. Верхняя граница рассматриваемой системы (ε0sat) реализуется при насыщающей интенсивности света (i0 =1), при интенсивности света выше этого предела соотношение ресурсные/структурные формы не будет изменяться, т.е. является максимальным:
Решение системы дает зависимость соотношения ресурсные/структурные формы биомассы микроводорослей от интенсивности света после ресурсного насыщения:
Окончательно зависимость соотношения ресурсные/структурные формы биомассы микроводорослей от интенсивности света в непрерывной культуре невысокой плотности записывается в виде двух уравнений:
Краткий анализ полученных уравнений и их графическое представление Определим экстремальные величины искомых соотношений, которые можно выразить через кинетические константы и экономические коэффициенты. Это позволит полученные зависимости выразить в безразмерных величинах. Максимальное значение соотношения ресурсные/структурные формы биомассы можно принять за нормирующий множитель и выражать полученные зависимости в долях от максимального значения:
Кинетические коэффициенты уравнений отнесем к максимальной удельной скорости преобразования ресурсной формы биомассы в структурные составляющие клетки. Учитывая, что максимальная скорость роста и фотосинтеза более чем на порядок выше удельной скорости поддержания, примем, что их отношение отличается в 20 раз. Экономический коэффициент qrs примем равным 40%, а isat = 0,5. Последняя величина накладывает ограничения:
Примем, что:
Полученные видоспецифические параметры отражены в таблице:
На рис. 1 и 2 показаны полученные зависимости при использовании именно таких коэффициентов и параметров микроводорослей.
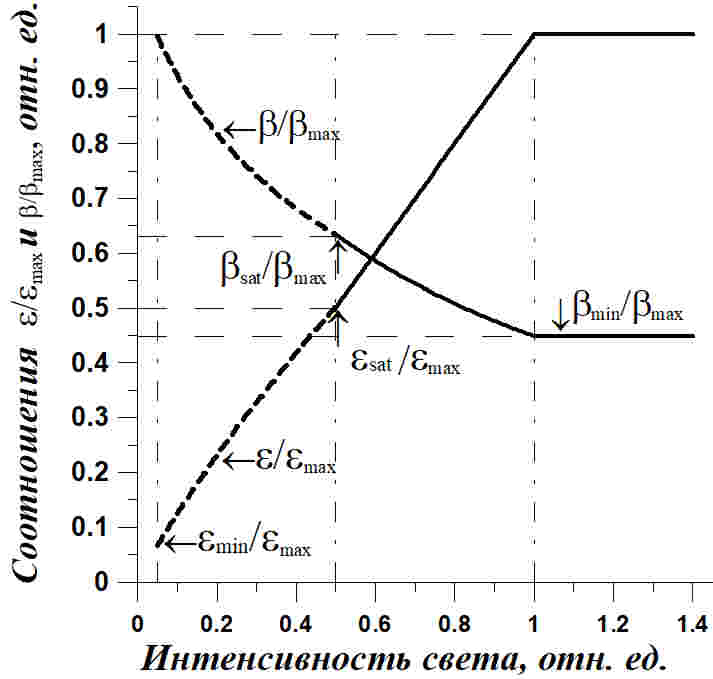
Рис. 1. Зависимость соотношения ресурсных и структурных форм (ε*=εmax) и содержания структурных компонентов в биомассе микроводорослей (β*/βmax) от интенсивности света в непрерывной культуре низкой плотности (isat = 0,5) Fig. 1. The dependence of the ratio of resource and structural forms (ε*=εmax) and content of structural components in the microalgae biomass (β*/βmax) on light intensity in continuous culture of low density (isat = 0,5)
Из представленного рисунка видно, что после ресурсного насыщения скорости роста микроводорослей соотношение резервные/структурные макромолекулярные формы биомассы клеток увеличивается с ростом интенсивности света вплоть до насыщающей. В данном случае соотношение увеличивается в 2 раза. В то же время содержание структурных форм в биомассе клеток уменьшается до минимальных значений. Это происходит за счет накопления в биомассе ресурсных форм. В реальности это должно приводить к увеличению углеводов или липидов в клетках при увеличении интенсивности действующего света, что и подтверждается многочисленными экспериментальными данными.
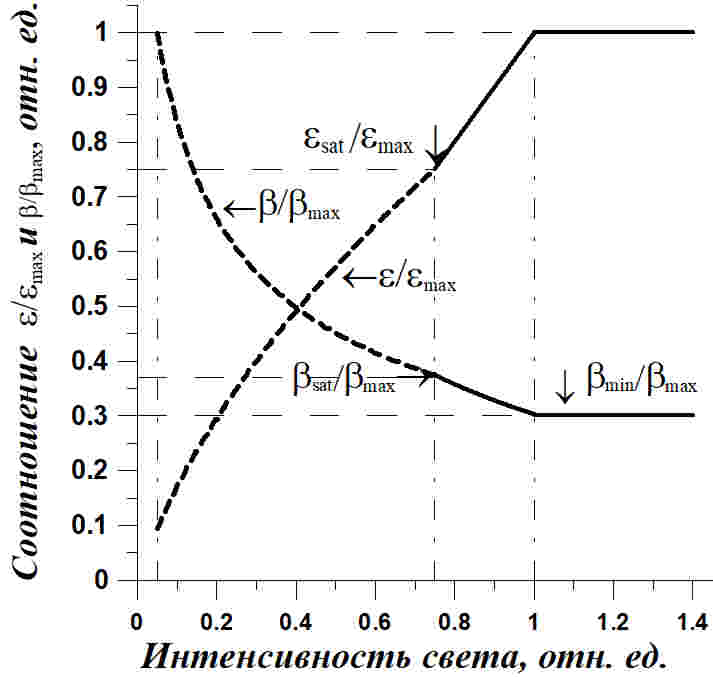
Рис. 2. Зависимость соотношения ресурсных и структурных форм (ε*=εmax) и содержания структурных компонентов в биомассе микроводорослей (β*/βmax) от интенсивности света в непрерывной культуре низкой плотности (isat = 0,75) Fig. 2. The dependence of the ratio of resource and structural forms (ε*=εmax) and content of structural components in the microalgae biomass (β*/βmax) on light intensity in continuous culture of low density (isat = 0,75)
На рис. 2 показаны эти же зависимости, но полученные для случая, когда ресурсное насыщение ниже светового на 25% (isat = 0,75). Интерес здесь представляет отношение максимального значения содержания структурных форм в биомассе в компенсационной точке к минимальному значению при световом насыщении. Считая, что эта величина косвенно отражает содержание белка в биомассе микроводорослей, можно сказать, что за счет света, при прочих не лимитирующих рост условиях, это содержание может изменяться более, чем в три раза.
Работа выполнена в рамках гос. задания № 1001-2014-0017 «Разработка научных основ решения гидробиологических и биотехнологических проблем интегрированного управления прибрежными зонами».
Список литературы 1. Ивлев А.А. Колебательная природа углеродного метаболизма в фотосинтезирующей клетке по данным изотопного состава углерода // Успехи современной биологии. 2011. № 2 (131). С. 178–192. 2. Минкевич И.Г. Математические проблемы организации метаболических путей из биохимических реакций // Математическая биология и биоинформатика. 2016. Т.11. №2. С. 406–425. doi: 10.17537/2016.11.406 3. Перт С. Дж. Основы культивирования микроорганизмов и клеток. – М., 1978. – 333 с. 4. Рубин А.Б. Биофизика Т.3. М.: ИКИ, 2013. – 480 с. 7. Физиология растений / под ред. проф. Ермакова И.П. — М.: Академия, 2005. – 640 с. 8. Blackman F.F. Optima and limiting factors // Ann. Bot. Lond. 1905. 19. P. 281–295. 9. Droop M.R. Some thoughts on nutrient limitation in algae // J. Phycolog. 1973. 9. P. 264–272. 10. Droop M.R. The definition of X and Q in the cell quota model // J. Exp. Mar. Biol. Ecol. 1979. 39. P. 203. 11. Liebig J. Chemistry in its Application to Agriculture and Physiology. – London: Taylor and Walton, 1847. – 320 p. Статья поступила в редакцию 15.12.2017
Influence of light on macromolecular composition of microalgae Rudolf P. Trenkenshu It is shown that the form of dependence of macromolecular composition of microalgae biomass on light is determined by the ratio of saturating values of photosynthesis and synthesis of structural components of cells. When processes rates are limited in the sequence of conversion of resource substances into structural ones, with no light saturation achieved, spare substances in the form of carbohydrates and possibly lipids are accumulated in the cells. The equations allowing to describe the dependence of ratio of resource (reserve) and structural forms of biomass on the light intensity in dynamically equilibrium (continuous) culture of low density are obtained. Key words: microalgae; cultivation; photosynthesis rate; specific growth rate; biochemical composition.
Об авторе Тренкеншу Рудольф Павлович – Trenkenshu Rudolf P. кандидат биологических наук r.trenkenshu@rambler.ru Корреспондентский адрес: Россия, 299011, Севастополь, пр. Нахимова, 2, ФГБУН ИМБИ; тел. (869)-2550795.
ССЫЛКА НА СТАТЬЮ: Тренкеншу Р.П. Влияние света на макромолекулярный состав микроводорослей в непрерывной культуре невысокой плотности (Часть 2) // Вопросы современной альгологии. 2017. № 3 (15). URL: http://algology.ru/1241 Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно.
При перепечатке ссылка на сайт обязательна
На ГЛАВНУЮ
|
|||
|
| ||