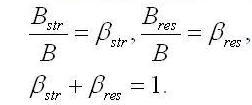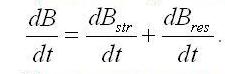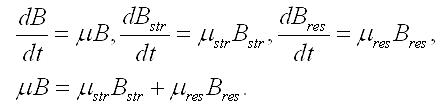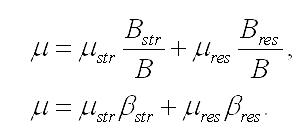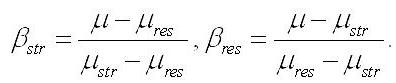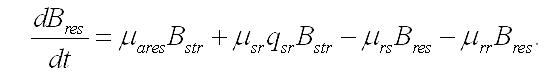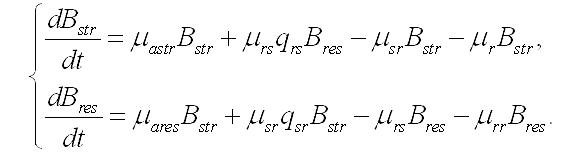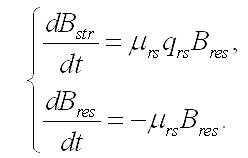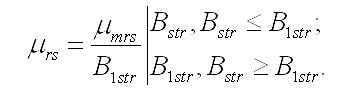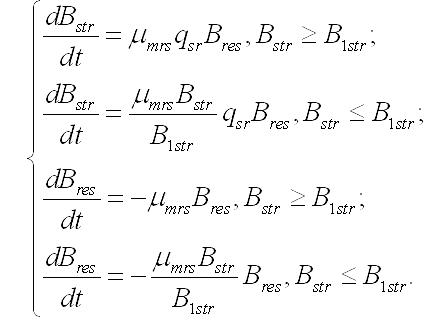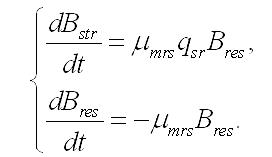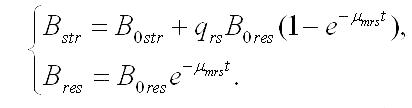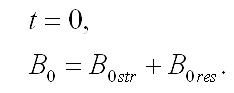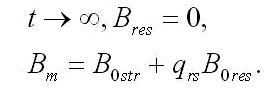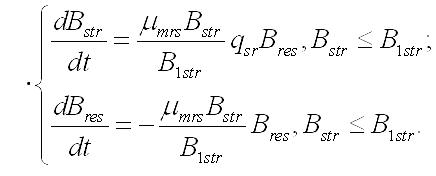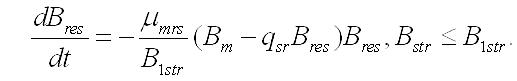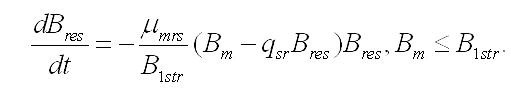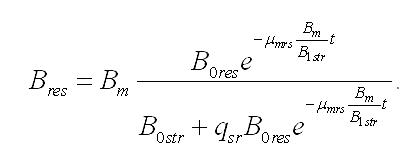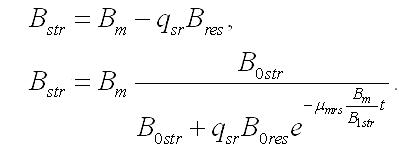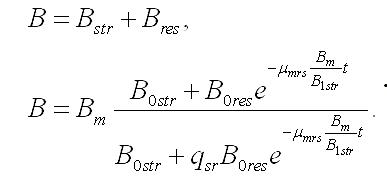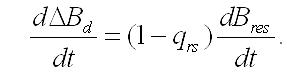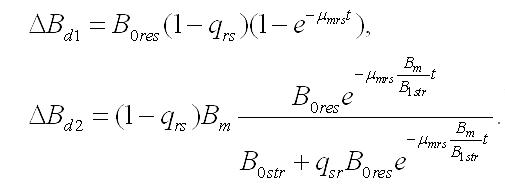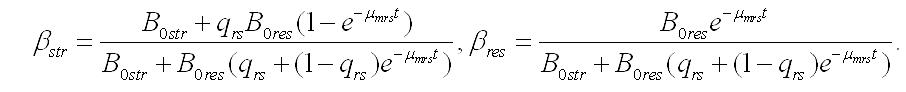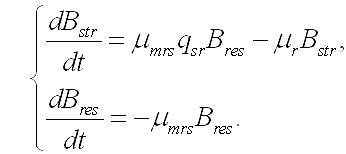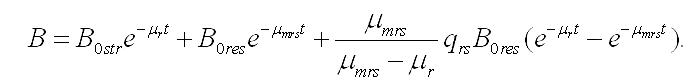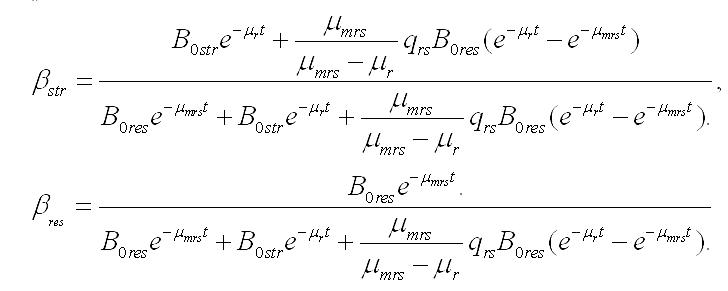|
|
|
Динамическая модель биотрансформации резервных и структурных форм биомассы микроводорослей в темноте
Тренкеншу Р.П. Rudolf P. Trenkenshu
Институт морских биологических исследований им. А.О. Ковалевского (г. Севастополь)
УДК 582.232:581.121
На основе представления о биомассе, как совокупности структурных и резервных форм, и кинетики темновых реакций фотобиосинтеза микроводорослей, предложена система уравнений, описывающая переходы резервных веществ в структурные компоненты клеток. Показано, что в случае, когда скорость трансформации резервной составляющей не зависит от уровня структурной компоненты, кинетика трансформации определяется только удельной скоростью превращения резервных компонент в структурные. Со временем биомасса будет представлена только структурной компонентой. Если процесс трансформации резервных компонент в структурные зависит от содержания последних в биомассе, то кинетика перехода одной формы в другую зависит также и от доли структурной компоненты. Со временем структурная компонента будет определять биомассу. Если на процессы превращения одной формы в другую влияет процесс окисления структурной компоненты, то темновая потеря биомассы определяется двумя процессами: более быстрым процессом превращения резервной формы в структурную и процессом деструкции структурных форм в результате темнового дыхания. При этом биомасса культуры постоянно снижается во времени. Ключевые слова: микроводоросли; культивирование; компоненты биомассы; темновые процессы; темновая потеря биомассы; моделирование.
Введение. Микроводоросли, как и другие растения, запасают энергию и углерод в форме фосфорилированных сахаров (цикл Кальвина), которые служат углеродными скелетами и энергетическим субстратом для синтеза всех прочих соединений. Таким образом, сахар, образующийся в процессе фотосинтеза из углекислоты, — это основное органическое вещество, которое обеспечивает энергией и пластическим материалом последующие темновые реакции в клетке, уже не зависящие от света. При дневном освещении происходит накопление продуктов цикла Кальвина, преимущественно в виде полисахаридов, которые используются для синтеза биомассы в ночное время. В любом случае темновые реакции происходят независимо от освещения и играют важнейшую роль для роста микроводорослей. При этом скорость роста микроводорослей определяется как структурными формами биомассы клеток в виде метаболических структур, так и текущей концентрацией свободных (в физиологии растений используются термины – резервные или запасные) веществ в клетке, которая зависит от соотношения скоростей их синтеза и расхода. Вместе с этим, изменение уровня резервных углеводов приводит к изменению общего биохимического состава клеток микроводорослей. Несмотря на большое количество работ по биохимическому составу микроводорослей и факторов, их определяющих, относительно мало модельных представлений, позволяющих объяснить и количественно описать полученные экспериментальные результаты. Особенно это касается темновых процессов. Предлагаемая работа представляет попытку количественно описать динамику двух основных составляющих биомассы микроводорослей, структурную и резервную, при помещении клеток в темноту.
Общий подход. Наиболее общее представление о темновых процессах в культурах микроводорослей связано с понятием о «темновой потере биомассы» в фотоавтотрофной культуре микроводорослей (Eppley, Sharp. 1975, Edmundson, Huesemann. 2015). Для гетеротрофных, миксотрофных и фотогетеротрофных культур применение этого понятия ограниченно (Perez-Garcia et al., 2011). Эта потеря в основном связана с окислением углеводов в клетках планктонных микроводорослей (Eppley, Sharp, 1975) или темновым дыханием микроводорослей в культуре (Grobbelaar, Soeder, 1985), скорость которого измеряется по скоростям поглощения кислорода и выделения СО2 (Eppley, Sharp, 1975; Grobbelaar, Soeder, 1985). Вместе с этим во время нахождения клеток в темноте кроме резкого снижения углеводов наблюдается накопление белков и некоторое снижение липидов в культуре микроводорослей (Hu et al., 1996). Иными словами, в темноте происходит как уменьшение биомассы за счет потери углеводов и липидов, так и её увеличение за счет синтеза белков и других клеточных составляющих. Т.е. темновые процессы представляют собой совокупность множества взаимосвязанных процессов, полный учет которых при математическом моделировании приведет к большому количеству уравнений или коэффициентов и бессмысленности моделирования в целом. Предельным упрощением для снижения количества уравнений без потери смысла явлений будет выделение двух основных величин, соотношение которых может служить характеристикой происходящих процессов. Такой подход использован в данной работе.
Структурные и резервные формы биомассы. Выделим в биомассе клеток основную часть, включающую все структурные компоненты клеток. Остальная часть будет представлять собой все вещества, запасенные в процессе роста клеток, включая интермедиаты, т.е. биомассу микроводорослей (B) разделим на две части: структурную (Bstr) и запасную (резервную, Bres).
Обозначим доли этих частей в биомассе:
Возможно, что часть биомассы будет представлять собой неструктурные инертные элементы, не участвующие в процессе биотрансформаций. Количество этих веществ, во-первых, можно считать малым, во-вторых, будет постоянным и не повлияет на скорости преобразований. В процессе метаболизма микроводорослей скорость синтеза биомассы будет определяться суммой скоростей синтеза структурных и накопления или расхода резервных составляющих:
Эти скорости можно выразить через удельные скорости (μ, μstr , μres) и текущие значения биомассы и её составляющих:
Отсюда находим связь удельной скорости синтеза биомассы с удельными скоростями синтеза или расхода составляющих:
Последнее уравнение показывает, что удельная скорость роста микроводорослей зависит не только от удельных скоростей синтеза отдельных составляющих, но и от их соотношения в клетках. С учетом того, что доли составляющих биомассы в сумме дают единицу, получим формулу связи долей данного компонента биомассы с удельными скоростями синтеза или деструкции биомассы и её составляющих:
Продукция составляющих. Скорость накопления (или убыли) структурных и резервных веществ в клетке зависит не только от внешних условий, но и от взаимных переходов одних форм в другие. Это касается как энерго-, так и массообмена. Особенно ярко это проявляется в темновой стадии фотосинтеза (цикле Кальвина), где из неорганического углерода образуются фосфорилированные сахара, являющиеся источником энергии и углеродных скелетов для всех последующих реакций синтеза. При фотоавтотрофном росте микроводорослей скорость изменения структурных форм биомассы будет зависеть от удельных скоростей: - световых стадий фотосинтеза, участвующих в синтезе структур (μastr), - трансформации резервных составляющих в структурные (μrs), - трансформации структурных составляющих в резервные (μsr), - необратимого распада структур и их экскреции (μr). При таких метаболических трансформациях необходимо также учитывать стехиометрию переходов. Этот фактор будем учитывать коэффициентом (q) с соответствующим подстрочным индексом. Динамика изменения структурных компонентов будет описываться следующим уравнением:
Скорость изменения резервных составляющих биомассы будет зависеть от удельных скоростей: - световых стадий фотосинтеза, участвующих в синтезе резервных веществ (μares), - трансформации резервных составляющих в структурные (μrs), - трансформации структурных составляющих в резервные (μsr), - экскреции запасных веществ во внешнюю среду (μrr):
В результате имеем систему уравнений, описывающую динамику структурных и резервных составляющих биомассы клеток при фотоавтотрофном росте:
Темновые процессы. Рассмотрим упрощенный вариант трансформации резервных веществ в структурные компоненты биомассы клеток микроводорослей. Такой вариант возможен при помещении клеток в темноту, при отсутствии внешнего органического питания и скоростях распада биомассы много меньших скорости перехода запасов в структуры. Система уравнений редуцируется до вида:
Рассмотрим коэффициенты, входящие в уравнения полученной системы. Коэффициент трансформации (qrs), показывающий количество единиц структурных веществ, которые получаются за счет расхода единицы запасных, можно считать величиной постоянной. Величина (qrsB0res) показывает, какое количество структурных компонентов можно дополнительно получить при переходе всех начальных запасов (B0res) в структуры. Фактически этот коэффициент представляет собой полный аналог экономического коэффициента – понятия, используемого в микробиологии Y (Перт, 1978). Коэффициент трансформации всегда будет меньше единицы за счет энергетических трат на синтез структур. Применительно к микроводорослям эти траты будут представлять собой расходы на дыхание (в основном, потери углерода в виде углекислого газа при окислении полисахаридов). Количественно потери биомассы в темновом процессе (темновая потеря биомассы, ТПБ) оцениваются по величине расхода резервных форм:
В оба уравнения системы входит удельная скорость биотрансформации резервных форм в структурные. Переход является ферментативным процессом, т.е. скорость будет зависеть от количества фермента (или транспортных структур), контролирующего процесс, и от субстрата, представляющего собой резервное вещество. Учитывая, что синтез структурных компонентов происходит в структурах клетки, необходимо принять прямую пропорциональность между количеством фермента и структурной биомассой. Удельная скорость биотрансформации будет максимальной (μmrs) и независимой от резервных веществ при их высоких концентрациях, превышающих некоторую величину (B1str), снижение концентрации ниже этой величины, приведет к пропорциональному снижению удельной скорости перехода (Тренкеншу, 2009):
В результате получим систему уравнений:
Динамика биомассы и её форм в культуре. 1. Наиболее простым представляется вариант системы, в которой удельная скорость биотрансформации зависит только от резервных веществ и не зависит от структурных, т.е. максимальна. Тогда система упростится:
Интегрирование уравнений при начальных условиях (при t=0, Bstr= B0str, Bres= B0res, B0str + B0res= B0) позволяет описать изменение составляющих микроводоросли со временем (при нахождении клеток в темноте):
Зависимость содержания биомассы микроводорослей в культуре от времени нахождения клеток в темноте, если считать удельную скорость биотрансформации резервных веществ в структурные постоянной величиной, описывается уравнением:
В начальный момент времени нахождения клеток в темноте:
После длительного содержания микроводорослей в темноте все резервные формы трансформируются в структурные, которые достигают максимального значения, равного минимальному значению биомассы в культуре:
С учетом минимального значения уравнение для динамики биомассы примет вид:
Динамика биомассы и её форм в культуре. 2. Если удельная скорость биотрансформации зависит не только от резервных веществ, но и от структурных, то система уравнений запишется в виде:
Разделив первое уравнение на второе, получим дифференциальную связь между структурными и резервными составляющими биомассы:
Интегрируя при начальных условиях, получим связь между формами биомассы:
Подставляя полученное выражение во второе уравнение системы, имеем:
Уравнение справедливо только для концентрации структурных форм ниже точки смены лимитирующих факторов,
В результате интегрирования этого уравнения при начальных условиях путем разделения переменных получим зависимость содержания резервных форм биомассы микроводорослей от времени их нахождения в темноте:
Изменения в содержании структурных форм биомассы в культуре находим из уравнения баланса:
Концентрация биомассы в культуре будет представлять собой сумму структурных и резервных составляющих: Обе представленные модели приводят к стабилизации концентрации биомассы в культуре микроводорослей после длительного содержания в темноте (Bm), причем биомасса представлена в основном структурными формами.
Темновая потеря биомассы (ТПБ). Скорость ТПБ будет пропорциональна скорости трансформации резервных составляющих в структурные:
Потери представляют собой разность между начальной биомассой микроводорослей и её текущим значением:
После длительного содержания в темноте величина ТПБ будет максимальной:
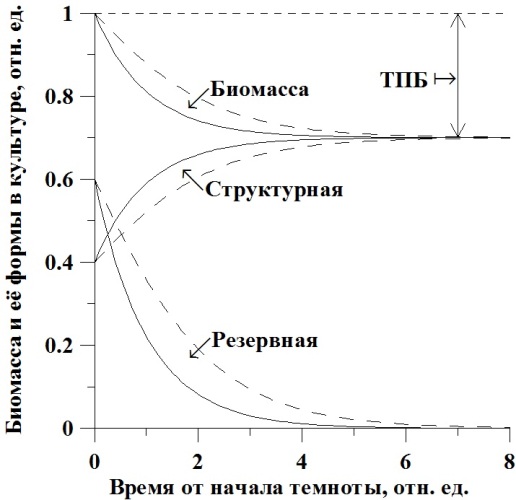
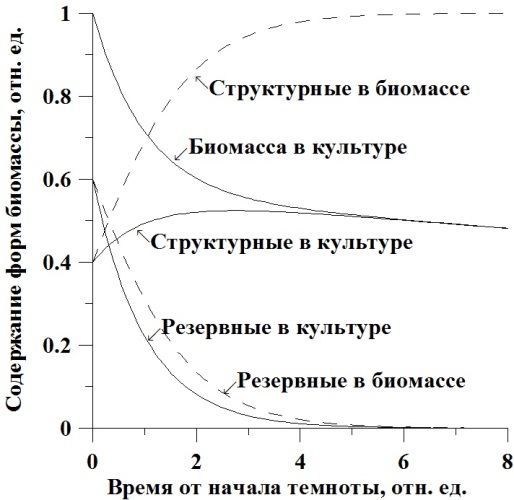
Полученные уравнения графически представлены на рис. 1. В качестве начальных данных в уравнениях принята концентрация биомассы, состоящая на 40% из структурных форм и на 60% из резервных. Коэффициент трансформации принят равным 50%. Шкала времени условная, выражена в единицах μmrst, μmrs=1. Сплошные линии – расчет для удельной скорости, не зависящей от структурных составляющих. Для зависимой скорости – штриховые линии.
Рис. 1. Динамика изменения концентрации биомассы и её структурных и резервных форм в культуре (пояснения в тексте).
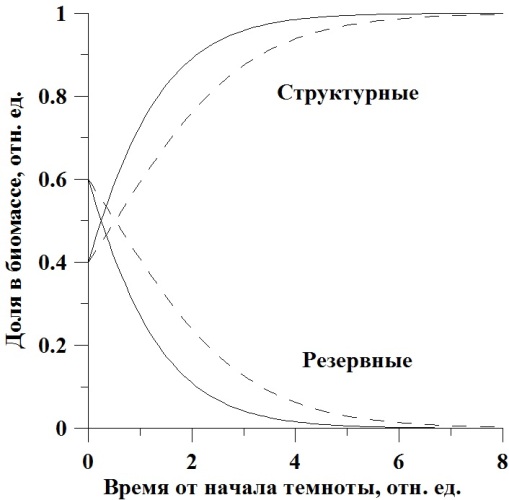
Динамика составляющих в биомассе. Иная картина наблюдается при оценке содержания резервных и структурных форм в биомассе микроводорослей при их помещении в темноту. Доля составляющих веществ при максимальной удельной скорости биотрансформации будет изменяться согласно уравнениям:
Зависимости, рассчитанные по этим уравнениям и с теми же начальными условиями и коэффициентами, показаны на рис. 2.
Рис. 2. Динамика изменения концентрации структурных и резервных форм в биомассе клеток микроводорослей (пояснения в тексте).
Динамика биотрансформаций при необратимом распаде структур. Более близкой к реальности можно рассматривать модель, учитывающую распад структурных компонентов биомассы. В микробиологии этот процесс рассматривают как расход энергии (в виде потери биомассы) на поддержание структур клетки, не связанный с расходом на рост. В этом случае система уравнений при максимальной скорости биотрансформации запишется в виде:
Второе уравнение системы имеет такой же интегральный вид, как и в предыдущих моделях:
Подставляя полученное выражение в первое уравнение, получим:
Интегрируя при начальных условиях, получим зависимость содержания структурных форм биомассы в культуре от времени содержания клеток в темноте:
Сумма структурных и резервных форм позволяет описать зависимость концентрации биомассы в культуре от времени:
Динамика содержания структурных и резервных форм в биомассе микроводорослей описывается уравнениями:
Полученные уравнения графически представлены на рис. 3. В качестве начальных данных в уравнениях приняты такие же значения, как на предыдущих рисунках, удельная скорость распада структурных форм в пятьдесят раз ниже удельной скорости биотрансформации.
Рис. 3. Динамика изменения концентрации структурных и резервных форм в культуре и биомассе клеток микроводорослей (пояснения в тексте).
Принципиальным отличием последней модели является неуклонное падение концентрации биомассы в культуре при длительном нахождении клеток в темноте.
Темновая потеря биомассы при необратимом распаде структур. Полученные зависимости позволяют найти динамические выражения для темновой потери биомассы микроводорослей в культуре и доли резервных и структурных форм в биомассе клеток. Динамика ТПБ в культуре будет описываться уравнением:
Из уравнения следует, что темновая потеря биомассы постоянно увеличивается со временем.
Описание экспериментальных данных. Необходимо отметить, что в литературе практически отсутствуют надежные измерения динамики темновой потери биомассы микроводорослей в культуре. Редким исключением являются результаты опытов, использованные нами в работе (Trenkenshu et al., 2013). B основу этой работы были взяты косвенные данные по динамике окисления углеводов в ночное время (Eppley, Sharp, 1975), ТПБ (Torzillo, 1991) и результаты измерения содержания белков, углеводов и липидов в культуре микроводорослей на закате и рассвете (Hu et al., 1996). Позднее А.Л. Авсиян (Avsiyan, 2014), для описания тех же динамических данных (Trenkenshu et al., 2013), предложила упрощенную эмпирическую модель, по форме соответствующую модели 1 в данной работе. К сожалению, точность измерений не позволяет сделать выбор в пользу той или иной модели.
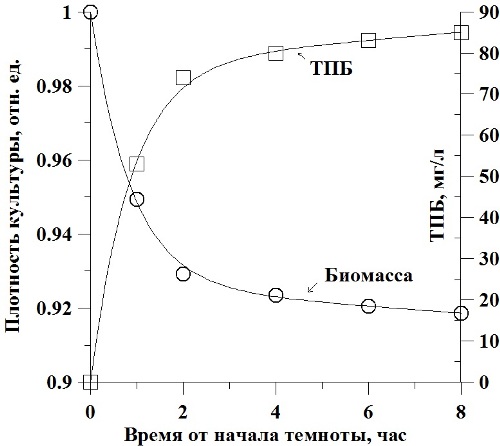
Рис. 4. Динамика изменения концентрации биомассы и темновой потери биомассы в культуре микроводорослей (пояснения в тексте).
На рис. 4 представлены экспериментальные точки и расчетные кривые динамики биомассы в культуре и темновая потеря биомассы спирулины для модели, учитывающей распад структурных форм. Показано хорошее совпадение модели с экспериментальными данными, однако необходимы дальнейшие более точные исследования, особенно по динамике биохимических трансформаций у микроводорослей в темноте.
Выводы: 1. Рассмотрена гипотеза, в которой биомасса представлена двумя компонентами: резервной (мобильной) и структурной (консервативной). Предложена общая система дифференциальных уравнений, описывающая динамику трансформации структурных и резервных составляющих биомассы клеток при фотоавтотрофном росте. 2. Показано, что в случае, когда скорость трансформации резервной составляющей не зависит от уровня структурной компоненты, кинетика трансформации определяется только удельной скоростью превращения резервных компонент в структурные. Со временем биомасса будет представлена только структурной компонентой. 3. Если процесс трансформации резервных компонент в структурные зависит от содержания последних в биомассе, то кинетика перехода одной формы в другую зависит также и от доли структурной компоненты. Со временем структурная компонента будет определять биомассу.
Список литературы 1. Перт С. Дж. Основы культивирования микроорганизмов и клеток. – М., 1978. – 333 с. 2.Тренкеншу Р.П. Культура микроводорослей как модельный объект в гидроэкологии // Морской экологический журнал. 2009. Т. 8, вып. 4. C. 41-52. 3. Avsiyan A.L. Dynamics of loss of biomass in culture of Arthrospira platensis (Nordst.) Geіtler (Cyanoprokaryota) in darkness // Algologia. 2014. 24(3). P. 417-420. 4. Edmundson S.J., Huesemann M H. The dark side of algae cultivation: Characterizing night biomass loss in three photosynthetic algae, Chlorella sorokiniana, Nannochloropsis salina and Picochlorum sp.// Algal Research. 2015. V. 12. P. 470-476. 5. Eppley R.W., Sharp J.I. Photosynthetic measurements in the central North Pacific: The dark loss of carbon in 24-h incubations // Limnol. Oceanogr. 1975. V. 20 (6). P. 981-987. 6. Grobbelaar J.U., Soeder C.J. Respiration losses in planktonic green algae cultivated in raceway ponds // J. Plankton Res. 1985. V.7 (4). P. 497-506. 7. Hu Q., Guterman H., Richmond A. Physiological characteristics of Spirulina platensis (Cyanobacteria) cultured at ultrahigh cell densities // J. Phycol. 1996. V.32. P. 1066-1073. 8. Markager S., Sand-Jensen K. Patterns of night-time respiration in a dense phytoplankton community under a natural light regime // J. Ecol. 1989. V.77. P. 49-61. 9. Ogbonna C., Tanaka H. Night biomass loss and changes in biochemical composition of cells during light/dark cyclic culture of Chlorella pyrenoidosa // J. of Fermentation and Bioengineering. 1996. V.82 (6). P. 558-564. doi:10.1016/S0922-338X(97)81252-4. 10. Perez-Garcia O., Escalante F.M.E., de-Bashan L. E., Bashan Y. Heterotrophic cultures of microalgae: Metabolism and potential products // Water Research. 2011.V.45. P. З1-36. DOI: 10.1016/j.watres.2010.08.037. 11. Torzillo G., Sacchi A., Materassi R., Richmond A. Effect of temperature on yield and night biomass loss in Spirulina platensis grown outdoors in tubular photobioreactors // J. of Applied Phycology. 1991. V.3. P. 103-109. 12. Trenkenshu, R. P., Avsiyan, A. L., Novikova T M. The simplest models of microalgae growth. 9. Dark processes // Mors'kyi Ekolohichnyi Zhurnal. 2013. 12 (2). P. 69-77. 13. van Wagenen J. M., De Francisci D., Angelidaki I. Comparison of mixotrophic to cyclic autotrophic / heterotrophic growth strategies to optimize productivity of Chlorella sorokiniana // J. of Applied Phycology. 2015. 27(5). P. 1775-1782. DOI:10.1007/s10811-014-0485-1. Статья поступила в редакцию 18.04.2016
The dynamic model of biotransformation of reserved and structural biomass form of microalgae in darkness Rudolf P. Trenkenshu On the basis of available information on biomass, as the combination of structural and reserve forms, and the kinetics of darkness reactions of microalgae photosynthesis, a dynamic system of equations describing biotransformation of reserve substances to structural components of cells has been developed. Integral solutions allow one to quantify the dynamics of night biomass loss and decrease of biomass concentration in the culture in the dark. Key words: microalgae; cultivation; biomass components; darkness processes; night biomass loss; modeling.
Об авторе Тренкеншу Рудольф Павлович – Trenkenshu Rudolf P. кандидат биологических наук r.trenkenshu@rambler.ru Корреспондентский адрес: Россия, 299011, Севастополь, пр. Нахимова, 2, ФГБУН ИМБИ; тел. (869)-2550795.
ССЫЛКА НА СТАТЬЮ: Тренкеншу Р.П. Динамическая модель биотрансформации резервных и структурных форм биомассы микроводорослей в темноте // Вопросы современной альгологии. 2016. № 2 (12). URL: http://algology.ru/967 Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно.
При перепечатке ссылка на сайт обязательна
На ГЛАВНУЮ
|
|||
|
| ||