|
|
|
|
Экологическая физиология водных фототрофных организмов Часть 1. Водные оксигенные фототрофы 1.3. Экологические группировки водных оксигенных фототрофов
Экологические группировки водорослей Водоросли встречаются повсеместно: в реках и в морях, на поверхности почвы и в ее толще, на деревьях и скалах, стенах домов и в мехе животных. Их находят даже в местах, не доступных для других организмов: на больших глубинах, в пещерах и гротах, в снегу и в горячих источниках. Способность водорослей адаптироваться к разнообразным внешним условиям, неприхотливость и физиологическая пластичность способствовали расселению их по всему земному шару. Практически невозможно назвать какую-либо географическую зону на Земле, где отсутствовали бы эти голофитные (растениеподобные) организмы. Водоросли, т.е. растущие в воде, как и любые другие растения, нуждается в воде. Однако потребности в ней неодинаковы для разных их видов. Одни из них могут жить только при полном погружении в водную среду, другим достаточно грунтовых вод или дождя, третьи довольствуются совсем небольшими ее количествами – росой, брызгами водопада или прибоя. В зависимости от совокупности различно действующих экологических условий (освещения, температуры, характера субстрата и его химического состава и т.п.) водоросли образуют многообразные группировки, сообщества, или ценозы. Для каждого сообщества свойствен более или менее постоянный видовой состав. В целом водоросли можно разбить на следующие основные группировки: нейстонные (фитонейстон); плейстонные (фитоплейстон); планктонные (фитопланктон); бентосные водоросли (фитобентос); водоросли, находящиеся в толще льда, покрывающего поверхность водоема (фитопагон, криофитон); водоросли горячих источников (термофитон); водоросли соленых материковых водоемов (галофитон); воздушно-наземные или аэрофильные водоросли (аэрофитон); почвенные водоросли (фитоэдафон); водоросли подвижных и пустынных песков (псаммофитон). Представители первых семи группировок (ценозов) – типичные обитатели водной среды. Водоросли других группировок могут затапливаться лишь периодически. Качественный состав альгоценозов в пресных и морских водах неодинаков. Общие виды в них достаточно редки. По видовому составу к пресноводным альгоценозам близки альгоценозы суши. К этим двум типам альгоценозов (пресноводным и суши) можно отнести по видовому составу и альгоценозы наземных водоемов (озер) с соленой, но не морской водой.
Планктонные водоросли. Планктон (от греч. planktós – блуждающий) включает живое население водной толщи, состоящее из мелких растительных (фитопланктон) и животных (зоопланктон) организмов. Фитопланктон составляют свободноживущие микроскопические водоросли. Это преимущественно одноклеточные и колониальные формы, неподвижные или недостаточно подвижные, чтобы преодолевать пассивное перемещение водных масс[15]. ____________________________ [15] Кроме перечисленных фито- и зоопланктона выделяют еще так называемый сапропланктон, куда входят бактерии, грибы, актиномицеты. Функции этих трофических группировок в водных экосистемах различны. Фитопланктонные организмы, или первичные продуценты, усваивая неорганические соединения, создают в воде органическое вещество. Зоопланктонные организмы, или первичные консументы, питаются фитопланктоном и в свою очередь служат пищей для вторичных консументов, например рыб. Сапропланктонные формы, или деструкторы, в процессе жизнедеятельности разлагают органическое вещество до простых соединений.
Фитопланктон обитает в водоемах самой разной природы и размеров: от морей и океанов до маленьких пересыхающих луж и канав. Типичное фитопланктонное сообщество характерно для крупных водоемов: морей, больших озер и медленно текущих рек. В мелких водоемах и особенно в реках с быстрым течением к типично планктонным водорослям примешивается очень большое количество донных обитателей. Состав планктона водоемов или даже одного водоема в разных его участках по сезонам не остается постоянным. В течение года в связи с изменениями физических и химических условий в водоеме одни группы водорослей сменяются другими, а в периоды массового развития нередко доминирующим становится вообще один вид. Глубина, на которой обнаруживаются планктонные водоросли, зависит от типа водоема, прозрачности воды и изменяется, как правило, от нескольких метров в пресных водах до 100 м и более в морях. Отдельные группы представителей фитопланктона отмечаются и на более значительных глубинах. Максимальное количество фитопланктона обычно приурочено не к самым поверхностным слоям, а к более глубоким, причем для многих организмов характерен свой глубинный оптимум обитания. Такая стратификация фитопланктона объясняется неодинаковым температурным режимом, а соответственно и химическим составом различных слоев водной среды. Поддержанию выраженной слоистости в распределения озерного планктона в летний период способствует и стратификация[16] воды, затрудняющая вертикальное ее перемешивание. ____________________________ [16] Стратификация – расслоение воды по температуре: более холодные воды занимают придонное положение. Более теплые расположены сверху.
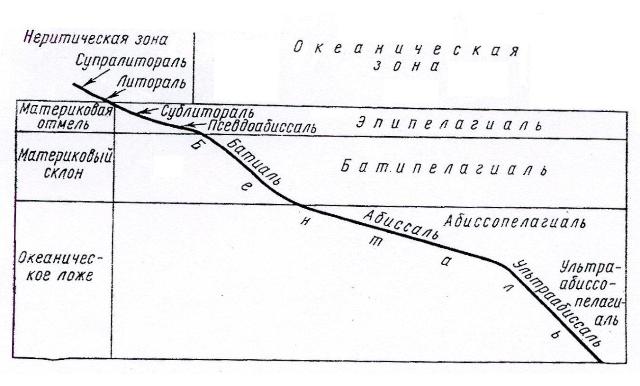
Пресноводный фитопланктон состоит главным образом из диатомовых, зеленых, синезеленых, золотистых, динофлагеллят и эвгленовых водорослей. Зеленые представлены монадными и коккоидными формами. Из них наиболее часто встречаются виды родов Chlamydomonas, Pandorina, Eudorina, Gonium, Volvox, Scenedesmus, Pediastrum, Ankistrodesmus, Kirchneriella, Oocystis, Chlorella и др. В заболоченных водоемах или в бассейнах, связанных с болотами, в планктоне преобладают десмидиевые водоросли: Cosmarium, Closterium, Staurastrum, Euastrum, Micrasterias, Xanthidium, Desmidium, Hyalotheca и др. Из синезеленых в пресноводном планктоне наиболее многочисленны Anabaena, Microcystis, Aphanizomenon, Gloeotrichia. Диатомовые представлены неподвижными формами из класса Pennatophyceae (Asterionella, Tabellaria, Fragillaria и т д.) и некоторыми видами из класса Centrophyceae (в основном различные виды рода Melosira). Видовое разнообразие типично планктонных диатомей в пресных водах невелико, но оно часто расширяется за счет временно планктонных видов, имеющих донную фазу в жизненном цикле. В сравнительно холодной воде нередко наблюдается изобилие монадных охрофит (виды родов Synura, Dinobryon, Uroglena, Mallomonas), в теплой воде – эвгленовых (виды родов Euglena, Trachelomonas, Phacus). Из динофлагеллят наиболее обычны виды Ceratium и Peridimium. Вышеперечисленные сообщества характерны в основном для средних и мелких по размеру пресноводных бассейнов: озер, прудов, заводей рек и т.д. В таких крупных озерах, как Байкал, Ладожское, Онежское, наблюдается много общего, особенно в распределении планктона, с морями. Melosira baicalensis (в Байкале) и M.islandia (в Онежском и Ладожском озерах), доминирующие в этих бассейнах, не опускаются на дно, как в более мелких водоемах, а образуют на некоторой глубине характерные зоны скопления, как в морях. Морской фитопланктон формируют в основном центричные диатомовые и динофлагелляты, синезеленые и кокколитофориды, зеленые водоросли отступают на второй план. Среди диатомей по видовому разнообразию наиболее многочисленны роды Chaetoceros, Thalassiosira, Rhizosolenia, Planktoniella и некоторые другие. За небольшим исключением (например, некоторые виды Rhizosolenia) это типично морские обитатели. Жгутиконосцы из отдела Dinophyta в морях представлены большим количеством видов, чем в пресных водах. Некоторые роды, такие как Dinophysis, Goniaulax, являются исключительно морскими обитателями. Кокколитофориды (известковые жгутиковые), несколько видов которых встречается и в пресной воде, в море представлены значительно разнообразнее. Только для морских бассейнов характерны силикофлагелляты (кремнежгутиковые водоросли). Число видов синезеленых водорослей несколько ограничено, тем не менее, один их вид – Oscillatoria (Trichodesmium) erythraea может развиваться так бурно, что вызывает «цветение» воды в тропических морях. В распределении морского фитопланктона четко прослеживается географическая зональность: в теплых тропических водах распространены преимущественно динофлагелляты и известковые жгутиковые, диатомовые же приурочены к холодным водам северных и арктических морей, где местами в больших количествах встречаются также силикофлагелляты и некоторые зеленые водоросли. В распределении фитопланктона в пределах одного бассейна (моря, океана) также существуют определенные закономерности. Это связано с делением поверхностных фотических[17] слоев водной толщи (или пелагиали) на 2 зоны: эпипелагиаль, или океаническую (глубоководную), существующую вне связи с грунтом, и неритопелагиаль, или неритическую (прибрежную), простирающуюся от берега до зоны конечных глубин фотической зоны, распологающейся на материковой отмели. В соответствии с этим весь планктон делят по месту его обитания на планктон эпипелагиали (океанический) и неритопелагиали (неритический). Распространение океанического фитопланктона также ограничено фотической зоной. Вода в этой области прозрачнее, чем в неритической зоне. Поэтому свет в ней распространяется на большую глубину. Однако эта зона более бедна биогенными элементами, особенно фосфатами. В результате продуктивность фитопланктона в ней несколько меньше, чем у берегов. Неритический фитопланктон отличается богатством видового состава и большей продуктивностью. По мере приближения к берегу пелагические формы здесь все больше соприкасаются с бентосными (рис. 3). ____________________________ [17] Фотический, или эвфотический слой – слой, в который проникает достаточное количество света, необходимое для протекания фотосинтеза; в среднем его глубина составляет 200-300 м.
Рис. 3. Экологические зоны бентали и пелагиали Мирового океана (по Константинов, 1986)
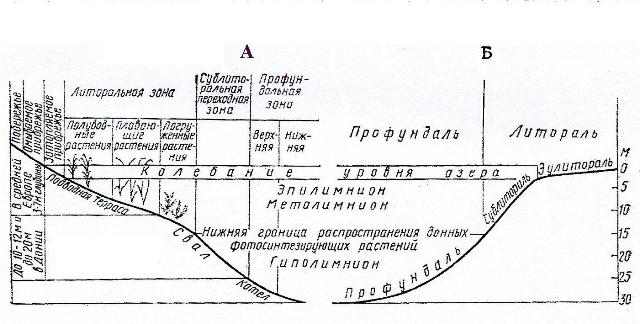
Фитопланктон опресненных внутренних морей заметно отличается по составу и по количеству видов не только от океанического, но и от неритического планктона Мирового океана (рис. 4). В открытой части этих водоемов, отличающихся пониженной соленостью по сравнению с океаном, отсутствуют типичные океанические виды. В неритическом планктоне встречаются лишь эвригалинные виды, а также виды, характерные для пресноводных водоемов, но приспособившиеся к условиям обитания в морской среде.
Рис. 4. Экологические зоны бентали и пелагиали озер: А – по Зернов, 1949; Б – по Ruthner, 1962.
Общая численность планктона в разных водоемах значительной колеблется. Например, в холодных арктических морях весной в 1 см3 верхних слоев воды насчитывается до 30 особей, в умеренных водах – до 100000 экземпляров и более. В пресноводных прогреваемых бассейнах это число резко возрастает. В водохранилищах при «цветении» воды (синезеленые водоросли) нередко в 1 см3 воды содержится до 1 млн. клеток. Иногда в прибрежных зонах таких «цветущих» водоемов скапливаются огромные массы водорослей, окрашивающих воду в ярко-зеленый или грязно-зеленый цвет. О биомассе фитопланктона того или иного водоема принято судить по количеству органического вещества, приходящегося на единицу поверхности или объема среды. В арктических морях она достигает 6-14 г/м3 воды, в Каспийском море – 1-3 г/м3, а недалеко от впадения в него Волги – 100-140 г/м3. В Азовском море летом биомасса иногда исчисляется в 250-300 г/м3. По аналогии с наземными растениями продуктивность (или урожайность) планктона оценивается количеством органического вещества, продуцируемого на 1 га водной поверхности за год. Годовая продукция планктона в пресных водах и морях выражается близкими величинами. Например, в Баренцевом море она равна 30-50 т (сырой массы) на 1 га, а в наиболее продуктивных пресных озерах – 26,5 т/ га. Предполагают, что биомасса фитопланктона в Мировом океане составляет примерно 1,5 млрд. т, а продукция его за год превышает биомассу в 300-400 раз, что возможно лишь при условии постоянного, ежесуточного деления фитопланктонных организмов, а соответственно возобновления их биомассы. Эти цифры как нельзя лучше подчеркивают значение планктонных водорослей как поставщиков органических соединений. Огромные водные пространства не являются бесплодными: благодаря развитию планктона урожаи с них не только не уступают, но иногда превосходят урожаи с наземных площадей. Планктонные водоросли, населяющие толщу воды, под действием силы тяжести неминуемо должны были бы опуститься на дно. Однако в природе этого не происходит, чему способствует как перемешивание воды, так и особенности в строении взвешенных организмов. Одной из этих особенностей является микроскопически малые размеры клеток этих форм. Кроме того, у некоторых водорослей, например монадных форм, этой же цели служат жгутики. Для увеличения плавучести есть и некоторые специальные приспособления, ведущие, во-первых, к уменьшению удельной плотности организмов и, во-вторых, к увеличению их трения о воду. Эти приспособления весьма сходны у планктонных водорослей разной систематической принадлежности, что свидетельствует о параллельном и независимом развитии их под влиянием одинаковых условий существования. Уменьшения удельной плотности организма достигается путем накопления в нем включений с плотностью меньше единицы. Это, например, капли жира, которые в качестве запасного продукта накапливаются в клетках фитопланктона; или наполненные воздухом газовые вакуоли, которые появляются у синезеленых водорослей, – они настолько уменьшают плотность клеток, что водоросли всплывают на поверхность воды и в периоды массового развития образуют там большие скопления. У планктонных диатомей «плавучесть» повышается за счет истончения, по сравнению с бентосными формами, панциря. Немаловажную роль играет и слизь, выделяемая многими планктонными водорослями. Для увеличения трения о воду у представителей родов Chaetoceros, Ceratium, Richteriella, Corethron, Stephanodiscus на стенке клеток имеются длинные шиповидные отростки, у Planktoniella, Ornithocercus – пластинчатые выросты, напоминающие парашют. Колониальные же представители фитопланктона с целью увеличения плавучести своего сообщества «придают» ему определенную форму: пластины у Pediasrum, плота у Scenedesmus, звездочки у Asterionella (ее радиально расположенные клетки иногда соединены нежными слизистыми нитями). У динофлагеллят также развиты приспособления, увеличивающие их поверхность, что также улучшает пассивную плавучесть клетки или колонии. Интересно отметить, что у водорослей, обитающих в теплых водах, приспособления к планктонному существованию более развитые, чем у холодноводных. Например, тропические представители Ceratium имеют более длинные роговидные выросты, чем представители этого же вида, обитающие в умеренном и холодном климате[18]. А у C. hirundinella, обитателя пресных вод, отмечен так называемый сезонный полиморфизм: в холодной воде (осенью и весной) этот вид имеет более короткие рога, чем в теплой воде (летом). Подобная температурно-сезонная изменчивость наблюдается и у представителей других групп. Причина этого явления – уменьшение вязкости воды с повышением ее температуры. Для миграции в толще воды ряд водорослей используют воздушные пузыри, образуемые ими в процессе метаболизма. ____________________________ [18] Существует также предположение, что выросты типа шипов и рогов сформировались как защитное устройство против выедания фитопланктона представителями зоопланктона. Нейстонные водоросли. Термином нейстон обозначено сообщество мелких растений и животных, живущих в зоне поверхностной пленки, у границы раздела водной и воздушной сред. В нейстоне выделяют две группы: эпинейстон, включающий организмы, живущие над поверхностной пленкой, и гипонейстон, в которую входят виды, прикрепляющиеся к поверхностной пленке снизу. Это сообщество организмов наиболее легко обнаружить в тихую погоду в мелких, защищенных от ветра водоемах (в лужах, торфяных карьерах, канавах, небольших заливах и прудах). Не исключается существование нейстона и в больших водоемах (в озерах и даже моря), но на сравнительно ограниченной площади спокойной воды (без волн). Обитатели поверхностной пленки нередко образую ярко окрашенный слой, видимый невооруженным глазом. Количество организмов на 1 мм2 поверхности в таких случаях составляет несколько десятков тысяч экземпляров. В пресноводном нейстоне наиболее многочисленны золотистые (виды Chromulina), эвгленовые (виды Euglena, Trachelomonas), зеленые (виды Chlamydomonas), разножгутиковые (виды Botrydiopsis) водоросли. Многие нейстонные организмы для удержания в зоне поверхностной пленки имеют специальные приспособления в виде слизистых колпачков, напоминающих маленькие парашюты, плавательных пластинок с гидрофобной поверхностью.
Плейстонные водоросли. Тело плейстонных водорослей отчасти находится в воде, а часть – над ее поверхностью. К типичным представителям фитоплейстона относятся, плавающие «маты» саргассовых водорослей. В которых находят убежища большое количество видов разных организмов.
Бентосные водоросли. К фитобентосу принадлежат все водоросли, жизнь которых так или иначе связана с дном водоема. Субстратом для них служат твердые и мягкие грунты (камни, скалы, песок, ил), другие растения (более крупные водоросли и высшие водные растения), панцири и раковины беспозвоночных животных и т.п., причем они могут либо прикрепляться к субстрату, либо свободно лежать на нем. В отличие от микроскопических планктонных организмов, бентосные водоросли представлены и микро-, и макроскопическими формами. Растут они, как правило, большими группами. Микроскопические формы в совокупности образуют видимые невооруженным глазом обрастания в виде слизистых пленок, войлочных подушечек, налетов, окрашенных в различные оттенки зеленого, желтого, бурого цвета. Макроскопические морские бентосные водоросли, некоторые виды которых достигают в длину 100 и более метров, формируют густые заросли, напоминающие подводные леса (например, заросли бурых водорослей). Для развития фитобентоса необходим прежде всего свет. Степень его использования, в свою очередь, зависит от сопряженного действия целого ряда других факторов: движения воды, температуры, характера грунта, содержания в воде минеральных и органических веществ и скорости поступления этих веществ в слоевище водорослей. Например, процессы фотосинтеза и дыхания, а также поглощение водорослями питательных веществ протекают более интенсивно в условиях постоянного обновления водных масс. Подтверждение этому – наиболее пышное развитие бентосных водорослей именно в местах с интенсивным перемешиванием воды: в реках и ручьях – на перекатах, где вода течет с большой скоростью, в морях – в проливах с течениями, в прибрежной прибойной зоне. Глубина обитания бентосных пресноводных водорослей составляет 30-40 м. Это связано с тем, что вода в большинстве пресных водоемов имеет низкую степень прозрачности. Дно глубоких бассейнов (в частности, озера Байкал) вообще свободно от фотоавтотрофных форм водорослей. В водоемах же с прозрачной водой эти растительные организмы опускаются на бóльшую глубину. Например, в озере Севан (Армения) Ulothrix, Cladophora, Enteromorpha вместе с диатомовыми растут на глубине до 70 м. Естественно, что в водоемах с разной степенью прозрачности воды и неодинаковым химическим составом не может быт идентичного распределения водорослевого бентоса по вертикали. Тем не менее, для пресных водоемов относительно закономерно, что верхний слой преимущественно заселяется особенно требовательными к свету зелеными водорослями. Глубже других в пресных водах опускаются диатомовые, т.к. некоторые их виды являются типичными сапротрофами, способными обитать на таких глубинах, куда не проникает свет. Пресноводный бентос состоит в основном из зеленых, разножгутиковых, диатомовых и синезеленых водорослей. Красные представлены небольшим числом видов, из них наиболее часто встречаются Batrachospermum. Среди зеленых водорослей довольно много прикрепленных видов, растущих на камнях и других твердых предметах (эпилиты), на высших растениях: тростниках, рдестах и т.п. (эпифиты). Наиболее распространены виды следующих родов: Ulothrix, Cladophora, Rhizoclonium, Stigeoclonium и т.д. Кладофора и стигеоклониум имеют вид ветвящихся кустиков. Иногда, отрываясь от субстрата, эти водоросли образуют в воде длинные «хвосты», одним концом прикрепленные к подводным растениям. Часто встречается также кустистая форма Draparnaldia. В прибрежной зоне оз. Байкал мощные заросли образуют эндемичные виды Draparnaldiella. В небольших водоемах (канавах, прудах) и в прибрежной области крупных озер обитает много неприкрепленных видов бентосных водорослей. Это разнообразные нитчатые формы зеленых (например, Spirogyra, Oedogonium), разножгутиковых (Tribonema) и синезеленых (Oscillatoria, Lyngbya, Tolypothrix) водорослей, а также диатомеи. Одни из них свободно лежат на песчаном или илистом дне (эпипелиты), другие располагаются на поверхности подводных растений, плотно прилегая к ним (эпифиты). Между высшими водными растениями можно обнаружить слизистые бесформенные скопления Tetraspora, мелкие округлые колонии Rivularia, Gloeotrichia pisum и др. Нитчатые водоросли образуют слизистую тину ярко-зеленого (зигнемовые водоросли) или буровато-зеленого (синезеленые водоросли) цвета. У некоторых видов днем нити, увлекаемые пузырьками кислорода, выделяемого при фотосинтезе, часто скапливаются большими массами на поверхности воды, а ночью снова оседают на дно. Миграция в толще воды с использованием воздушных пузырей характерна также для диатомовых водорослей. Самые крупные представители пресноводного фитобентоса – харовые водоросли. Виды хары и нителлы наиболее часто встречаются в лиманах, прудах и озерах с илистым дном, где они образуют густые заросли, прочно закрепляясь в субстрате с помощью длинных ризоидов. Макроскопический таллом до 1,5 м в длину имеет во взрослом состоянии и водяная сеточка (Hydrodictyon reticulatum). Для разрастания этой формы благоприятны прибрежные части прудов, ручьи, заводи рек, имеющие достаточно азотсодержащих солей. Из бентосных синезеленых водорослей по величине колоний, иногда достигающих размера куриного яйца, первенствует Nostoc pruniforme. В период массового развития скопления ностока могут препятствовать работе шлюзов и т.д. По внешнему виду морские бентосные водоросли сильно отличаются от нежных пресноводных. Их огромные заросли состоят главным образом из макроскопических бурых, красных и зеленых водорослей. В «подлеске» между крупными формами и эпифитно на них расселяются мелкие бурые и красные водоросли, которые, в свою очередь, могут обрастать разнообразными формами зеленых, диатомовых и синезеленых водорослей. Существует определенная связь между величиной таллома водорослей и характером грунта – размером частиц, его слагающих. Крупные водоросли обычно приурочены к каменистому и скальному грунту, обитают на сваях, подвижных субстратах (лодках, плотах и т.п.) и высших водных растениях. Рыхлый песок, мелкий галечник, ил, непригодные для макроскопических растений, заселяются обычно мелкими формами. Причина этого заключается в особенностях строения органов прикрепления макроскопических водорослей – короткие ризоиды или подошвы не проникают вглубь субстрата, а распластываются по его поверхности, для чего необходим плотный субстрат и относительно большая его поверхность. Длинные ризоиды, напоминающие корни высших растений и уходящие в песок или ил, у морских бентосных водорослей, так же как у пресноводных, встречаются у очень немногих форм. К ним относятся обитающие в пресных и соленых водах виды Chara и Nitella, а также виды Caulerpa, растущие на песчаном грунте в тропических морях. Флористический состав морского бентоса зависит от состава воды, обусловленного свойствами грунта и интенсивностью перемешивания водных масс, а также ее прозрачностью и температурой. Бентосные водоросли занимают прибрежную, сравнительно узкую часть моря. Глубина их обитания в разных морях неодинакова. Например, в северных морях России граница встречаемости бентосных растений проходит на незначительных глубинах – около 40-50 м, а у берегов Флориды она опускается до 100 м; в Средиземном же море, отличающемся необычной прозрачностью, бентосные водоросли растут на глубине 130-180 и более метров. Вертикальное распределение водорослей обусловлено особенностями прибрежной области, в которой они обитают (рис. 3). Эта область подразделяется на три зоны: супралитораль, литораль и сублитораль. Супралитораль – самая узкая полоса морского побережья, располагается выше линии наибольшего стояния воды во время приливов и орошается только брызгами прибоя. Литораль, или осушная зона, занимает пространство от высшего уровня прилива[19] до низшего уровня отлива. Один или два раза в сутки водоросли этой зоны остаются по несколько часов на воздухе[20]. Сублитораль, или инфралиторальная зона, находится ниже литорали и простирается до глубины эуфотической зоны. В ней выделяют две области: верхнюю, отличающуюся от литорали более спокойной водой и незначительной интенсивностью освещения, и нижнюю, наиболее глубокую часть прибрежной зоны, в которой растут водоросли. Она характеризуется наибольшим постоянством факторов среды и малым освещением. Альгофлора супралиторали в связи с нерегулярностью увлажнения и колебаниями в солености отличается сравнительно малым видовым разнообразием и представлена преимущественно микроскопическими формами. В литорали и верхней сублиторали обитают макроскопические бурые, зеленые и красные водоросли, образующие огромные заросли. В самых глубоких сублиторальных слоях на больших глубинах преобладают красные корковые водоросли[21]. ____________________________ [19] Высота приливов может быть очень разной: от нескольких сантиметров во внутренних морях до 10 м в открытых морях. [20] Береговая линия литоральной зоны может иметь разную протяженность: от нескольких метров до нескольких километров. Когда литораль достаточно обширна, ее подразделяют на горизонты: верхний, средний и нижний. [21] Преобладание красных водорослей обусловлено наличием у них фикобилиновых пигментов, способных поглощать в сине-зеленых частях спектра.
Широтное распределение морских бентосных водорослей обусловлено главным образом температурным фактором. Бурые макрофиты являются, как правило, типичными обитателями холодного и умеренного поясов (хотя, например, виды рода саргассум и диктиотовые характерны и для тропических, и для субтропических морей). Красные и зеленые водоросли преобладают в теплых морях. Широтное распределение бентосных водорослей можно проследить на примере Белого и Черного морей. Литоральную зону холодного Белого моря населяют в основном фукусовые водоросли (фукус, аскофиллум, пельвеция). Местами они занимают до 75% общей площади литоральных зарослей. Пельвеция желобчатая (Pelvetia canaliculata), самая мелкая из порядка Fucales, встречается в супралиторали и в верхней литорали. Фукус пузырчатый (Fucus vesiculosus) образует обильные разрастания в верхней и средней литорали. Далее следуют фукус двухрядный (F. destichus), аскофиллум узловатый (Ascophyllum nodosum), фукус зубчатый (F. serratus). Между крупными водорослями и отчасти на них поселяются более мелкие формы бурых (например, Chordaria flagelliformis ), а также красные (из родов Rhodymenia, Porphyra, Ceramium и др.) и зеленые (из родов Enteromorpha, Monostroma, Ulvaria, Ulva, Cladophora). Во время отлива эти водоросли оказываются защищенными от высыхания талломами крупных бурых водорослей. В верхней сублиторали густо разрастаются наиболее крупные ламинариевые водоросли (виды родов Laminaria, Chorda, Alaria и др.). На них много эпифитов, главным образом из красных водорослей: представителей родов Polysiphonia, Delesseria, Phycodris. Последние встречаются и на каменном грунте вместе с Phyllophora, Ahnfeltia и др. Для нижней сублиторали особенно характерны известновые красные водоросли, местами покрывающие дно сплошным ковром (виды рода Lithothamnion, Corallina). В теплых водах Черного моря преобладают красные водоросли, а бурые отступают на второй план. Здесь совсем нет ламинариевых, фукусовые представлены только цистозейровыми водорослями, однако у берегов Крыма и Кавказа они формируют плотные и обширные заросли: на глубине 10 м и более растет филлофора, образующая целые поля (филлофорное поле С.А.Зернова расположено в северо-западной части Черного моря; в настоящее время оно сильно уменьшилось). Для опресненных и загрязненных участков прибрежной зоны характерно массовое развитие зеленых водорослей (виды Cladophora, Enteromorpha, Ulva). Типичные для тропических морей сифоновые зеленые водоросли представлены здесь в основном кодиумом червеобразным (Codium vermiculare) и видами бриопсиса (Bryopsis). На распределение водорослей влияет также соленость: в бассейнах с океанической или близкой к ней соленостью альгофлора значительно богаче, чем в морях с низкими значениями этой характеристики.
Поскольку
донные водоросли обитают в сравнительно неширокой прибрежной полосе, не
превышающей одной десятой всей площади Мирового океана, суммарная биомасса этих
растительных организмов в целом уступает планктону, хотя в
отдельно взятых
конкретных точках она существенно превышает ее и колеблется от 0,5 до 60 кг/м2.
Водоросли горячих источников. Водоросли термофитона способны жить и размножаться в таких местообитаниях, которые на первый взгляд кажутся совершенно непригодными для жизни: в горячих минеральных источниках, загрязненных горячих водах, спускаемых фабриками и заводами. Температура воды в них часто намного превышает те границы (от 0 до 500С), в пределах которых живет большинство организмов. Водоросли найдены даже в воде с температурой, приближающейся к точке кипения. Термальным природным водам свойственна не только высокая, но и постоянная в течение года температура, обеспечивающая круглогодичную вегетацию водорослей. По химическим свойствам эти воды отличаются и высоким содержанием минеральных веществ и газа. Минеральный состав водной среды оказывает на флору часто не менее существенное влияние, чем температура[22]. ____________________________ [22] Поскольку водоросли очень чувствительны к химическому составу термальных вод, они могут служить индикаторами их качества (по составу водорослей судят о химических свойствах воды).
В пределах одного и того же горячего источника поддерживается стабильность жизненных условий, что обусловливает неизменность состава растений не только в течение года, но и на протяжении более длительного отрезка времени. Так, например, в ряде горячих источниках в настоящее время отмечается та же самая альгофлора, что и в прошлом столетии. Способность переносить экстремальные условия в течение всей жизни свойственна в основном синезеленым водорослям. Термофильных диатомовых и зеленых водорослей гораздо меньше. Например, в одном из горячих источников Камчатки из 52 обнаруженных видов водорослей на долю синезеленых пришлось более половины – 28 видов, диатомовых – 17, зеленых – только 7. Предел, при котором еще могут жить диатомовые и зеленые водоросли, не превышает 50,70С, тогда как у нитчатых синезеленых (из порядка Oscillatoriales) он равен 85,20С. В таких условиях синезеленые нитчатки образуют хорошо заметные пленки, плавающие на поверхности воды или лежащие на дне водоема. Общее количество видов водорослей, обнаруженных в горячих водах, – более 200. С повышением температуры в водоеме число видов, способных переносить изменение условий, заметно уменьшается. Большинство термофильных водорослей живет при температуре 35-400С. При 85-900С обнаружены единицы. Большая часть обитателей термальных вод относится к эвритермным организмам, живущим и при более низких температурах (роды Ulothrix, Oedogonium, Cladophora, Spirogyra, Cosmarium, Rhizoclonium, Nitzschia и т.д.). Специфических термофилов, не способных существовать при температуре ниже 300С, очень немного. Самые типичные и широко распространенные из них – Astigocladus laminosus и Phormidium laminosum. Оптимум их температурного развития лежит в пределах от 45 до 550С. У термофильные водорослей отсутствуют какие-либо специальными внешними приспособления, компенсирующие экстремальные воздействия среды, и поэтому морфологически не отличаются от форм, населяющих обычные холодные воды. Выносливость их, по-видимому, обусловлена внутренними физиологическими перестройками в клетках, структурой и химическим составом цитоплазмы.
Водоросли снега и льда. Представители этой группировки, приспособлены к жизни в крайних температурных условиях (при 00С и ниже). Фитопагон, криофитон, или по- другому, криофильные водоросли, обитают на поверхности снега и льда, а также их толще. В настоящее время список этих водорослей насчитывает свыше 100 видов. Среди них особенно распространены одноклеточные зеленые, диатомовые и синезеленые водоросли. Ограниченное число видов имеют золотистые водоросли и динофлагелляты. Водоросли пагона (от греч. pagos – лед и on – сущее) находятся в толще льда, покрывающего поверхность водоема. Внутри льда, особенно вблизи границы с водой, температура не опускается ниже одного градуса, что обеспечивает сохранение жизнеспособности организмов даже вмерзших в лед. В средних широтах Северного полушария в составе пагона обнаруживается до 150 видов различных организмов из различных таксономических групп. Впервые пагон был описан С.А. Зерновым. «Снежные» водоросли развиваются, как правило, не на свежевыпавшем, а на старом снегу, оставшемся лежать в холодных ущельях или на снежных полях высоко в горах. Их рост и размножение начинается после того, как под действием солнечной радиации снег частично растает. Окраска снега в зависимости от преобладания в нем того или иного вида (а также стадии развития) может быть красной, зеленой, желтой, бурой, почти черной. Наиболее известно явление так называемого «красного снега». Чаще всего оно связано с интенсивным размножением водоросли Chlamydomonas nivalis, в клетках которой содержится большое количество красного пигмента астаксинтина. Другой вид C. flavovirens, встречающийся в Высоких Татрах, окрашивает снег в желто-зеленый цвет. Позеленение снега в Гренландии, в Альпах и ряде других высокогорий обусловлено кроме видов Chlamydomonas видами Raphidonema, некоторыми десмидиевыми, синезелеными водорослями и динофлагеллятами. Буровато-желтую окраску обычно придают снегу диатомовые. Площадь окрашенного снега может занимать до нескольких квадратных километров. Во льдах арктических и антарктических бассейнов сильно разрастаются диатомеи. Первоначально они развиваются на нижней, погруженной в воду поверхности льда, а затем в толще его, распространяясь по тонким канальцам, которые пронизывают лед в разных направлениях. В случае массового развития диатомовые водоросли вызывают интенсивное буровато-желтое окрашивание льда на больших пространствах. При этом биомасса диатомей весьма значительна: с 1 м3 льда собирают около 1 кг (сырой массы) водорослей.
Водоросли соленых материковых водоемов. Своеобразную группировку организмов представляют собой водоросли соленых озер[23]. Концентрация солей в континентальных водоемах может быть очень высокой – до 285 г/л воды, если в ней преобладает хлористый натрий, и до 347 г/л при преобладании сульфата натрия. Солевыносливых водорослей довольно много, известны даже виды, живущие в насыщенных растворах. Альгофлора засоленных водоемов не может считаться морской, так как типичные обитатели моря здесь отсутствуют. По происхождению это, скорее, пресноводные формы, приспособившиеся к повышенной концентрации солей. Организмы, населяющие воды с хлористыми солями, делятся на две группы: эвгалофиты и олигогалофиты. К первой относятся формы, живущие только в присутствии солей натрия и магния (из них полигалобионты населяют воды с высокой их концентрацией, близкой к морской, а иногда и большей, а мезогалобионты – с уменьшенным содержанием солей, например во внутренних морях и в опресненных морских бухтах). Вторая группа включает формы, которые живут в воде либо без хлоридов (галофобы), либо с незначительным их содержанием, не более 5‰. Типичный полигалобионт – Dunaliella salina – одна из наиболее распространенных водорослей соленых водоемов, сходная по строению с хламидомонадой. В клетках этого вида дуналиеллы содержится красный пигмент b-каротин. В периоды массового развития пигмент из отмерших клеток водорослей переходит в солевой раствор (рапу), окрашивая его и соль, выпадающую в виде кристаллов, в разные оттенки красного цвета[24]. Другой обычный обитатель соленых озер – синезеленая водоросль Chlorogloea sarcinoides, встречающаяся на юге Украины. Мощные слизистые скопления этой водоросли образуют пласты в береговой части некоторых озер. Ил, остающийся после отмирания Chlorogloea, используется в лечебных целях. ______________________________ [23] Таких водоемов особенно много в районах с жарким климатом и там, где подстилающие породы содержат большое количество солей. [24] Рапа, в которой растет Dunaliella salina, имеет приятный запах фиалок
Типы симбиотических связей водорослей с другими организмами. Водоросли не всегда ведут свободный образ жизни. Очень часто можно наблюдать их сожительство – симбиоз – с бактериями, грибами, другими водорослями, высшими растениями и животными. Взаимоотношения между организмами, составляющими такие сочетания или комплексы, степень влияния одного компонента на другой могут быть самыми разными и не всегда носят характер положительных взаимодействий (когда один из видов извлекает для себя пользу из сожительства с другим видом и не причиняет ему никакого вреда). Обычно различают несколько типов сожительства водорослей с другими организмами: эпифитизм, эндофитизм, паразитизм, мутуализм. Эпифитизм представляет такой тип сожительства, при котором имеет место тесный наружный контакт между водорослью и другим организмом, на котором она поселяется, при автономности их питания. При эндофитизме автономность питания обоих организмов сохраняется, но между ними формируется уже внутренний контакт (водоросль живет внутри другого организма, не нанося ему видимого вреда). Паразитизмом называют такие взаимоотношения, когда внедрившаяся в тело хозяина водоросль питается за его счет и в конечном итоге приводит к отмиранию отдельных частей организма хозяина или иногда к полной его гибели. Мутуализм, наоборот, представляет собой такой тип взаимоотношений, когда совместно существующие виды (один организм внутри другого) извлекают из своего сожительства множество преимуществ и иногда не в состоянии жить раздельно. В природе такие случаи встречаются, правда, очень редко. Следует подчеркнуть, что резко разграничить перечисленные типы взаимоотношений, обнаружить их в чистом виде на практике удается далеко не всегда, скорее можно наблюдать переходные формы. Эпифитизм среди водорослей – очень частое явление. Он заключается в том, что водоросли-эпибионты используют другие растения как субстрат для прикрепления и, не проникая внутрь их тела, питаются вполне самостоятельно. При этом один и тот же вид водоросли может прикрепляться не только к разным растениям, но даже к неживым субстратам и животным[25]. В пресных водоемах эпифитами, например, являются диатомеи, поселяющиеся на талломе одной из распространенных зеленых водорослей – кладофоры, а в морях – многие бурые и красные водоросли, прикрепляющиеся к многолетним черешкам ламинарий, основаниям фукусов и других крупных форм. Все отделы водорослей имеют представителей, ведущих эпифитный образ жизни[26]. Приспособления к прикреплению у эпифитов разной систематической принадлежности часто носят конвергентный характер. Пример тому – удивительно похожие морфологически, но относящиеся к разным отделам коккоидные водоросли. ___________________________ [25] При использовании животного организма в качестве
места прикрепления водоросли (эпизоонты), как правило, тяготеют к
определенной группе животных.
[26] Необходимо заметить, что продолжительность эпифитирования у разных видов неодинакова: у одних она занимает почти весь жизненный цикл, у других – только часть его.
Отношения между эпибионтом и хозяином не всегда складываются безразлично для организма, служащего опорой. Это особенно наглядно проявляется при массовом развитии эпифитов, когда они почти сплошным слоем обрастают растение. Та же кладоформ или аквариумные высшие растения, плотно покрытые талломами эпифитных водорослей, испытывают явное угнетение. Причиной, по-видимому, служит избыточное затенение, нарушающее фотосинтез этих растений. Более специализированным и сложным типом сожительства двух организмов оказывается эндофитизм (эндосимбиоз), когда водоросль живет полностью или частично внутри другого организма. При этом хозяин не обнаруживает признаков какого-нибудь подавления, а эндофит, используя его тело как жизненное пространство и сохраняя способность самостоятельно питаться, не теряет своей независимости. Эндофит поселяется либо в толще клеточной стенки другой водоросли, либо проникает между клетками, раздвигая их. Например, Coleochaete nitellarum локализуется в клеточных стенках междоузлий нителлы, Endoderma - в стенках кладофоры. Водоросли родов Anabaena и Nostoc живут в воздушных камерах мхов Anthoceros и Blasia, Anabaena azollae – в полостях на нижней стороне листьев водного папоротника Azolla americana, а Nostoc punctiforme обитает в межклетниках растений Cycas и Zamia. Филлобиум (Phyllobium) поселяется вблизи сосудистых пучков некоторых высших растений (из разных семейств). Трентеполия (Trentepohlia) рассматривается одновременно и как эпифит, и как эндофит, поскольку глубоко проникает внутрь пробковой ткани коры деревьев. Цефалеурос (Cephaleuros) выступает как промежуточная форма между эндофитами и паразитами. Разрастаясь внутри живых листьев тропических растений, он вызывает отмирание прилегающих к нему тканей растения-хозяина. Однако сохраняя хлорофилл, он сохраняет способность к автотрофному питанию. Родохитриум (Rhodochytrium), встречающийся на сложноцветных, – уже настоящий паразит, живущий исключительно за счет хозяина, так как из–за отсутствия хлорофилла утерял способность к автотрофному питанию. Из красных водорослей наиболее совершенный пример паразитизма представляет лишенная хлорофилла Harveyella mirabilis, развивающаяся на красных водорослях Rhodomela. В отличие от вышеперечисленных примеров паразитизма эндосимбиозы могут характеризоваться и взаимовыгодными отношениями партнеров, наиболее тесно и длительно связанных друг с другом в едином комплексе. Классическим примером межклеточного эндосимбиоза служат лишайники, представляющие собой настолько прочное и гармоничное биологическое единство водоросли и гриба, что образуется целостный организм нового типа. В такой кооперации гриб защищает водоросль, а водоросль его кормит, так как сохраняет хлорофилл и способность к фотосинтезу. Правда, не всегда взаимоотношения между грибом и водорослью в лишайнике столь просты и безобидны, особенно для водоросли. Большая часть выгод от такого сожительства находится все-таки на стороне гриба, доминирующего в лишайниковых талломах. Внутриклеточный эндосимбиоз распространен значительно шире. В качестве симбионтов водоросли могут соединяться с животными – одноклеточными и многоклеточными (инфузориями, радиоляриями, гидрами, губками, некоторыми червями и др.). Сами они принадлежат чаще всего к простейшим зеленым водорослям и динофлагеллятам и носят специальное название зоохлорелла (Zoochlorella) и зооксантелла (Zooxanthella). В организмы одних видов животных водоросли попадают случайно, с пищей (благодаря голозойному типу питания). Внутри животного некоторые их клетки, которых почему-либо не коснулось переваривание, успевают приспособиться к необычным условиям существования и даже размножаются. В процессе жизнедеятельности они снабжают своего хозяина углеводами. Примерами таких в конечном итоге распадающихся симбиозов служат системы инфузории – зоохлореллы; личинки отдельных видов стрекоз – клетки Euglena gracilis, содержащиеся в эпителии пищеварительного тракта стрекоз; ресничный червь конволюта (Convoluta roscoffensis) – картерия (Carteria ) и др. Среди симбиотических пар встречаются и более прочные и продолжительные связи. Однако симбиотические отношения такого рода складываются у водорослей только с определенными видами животных. При этом автотрофный организм при размножении животных передается из поколения в поколение. Пример такого рода симбиозов – зеленая гидра-водоросль, асцидия-прохлорофитовая водоросль. Аналогичные симбионты имеются у Geosiphon и Rhizosolenia. Разница с предыдущими примерами в том, что здесь синезеленые водоросли свой облик заметно не меняют и их легко узнать. Это нитчатые формы, близкие к Nostoc и Anabaena. Geosiphon встречается на влажной почве и внешне напоминает маленькие ботридиумы. Таллом его состоит из стелющихся разветвленных нитей (без перегородок) и не отходящих от них вверх пузыревидных вздутий, содержащих нити ностока. В таком сообществе отсутствие пластид в растении-хозяине компенсируется за счет фотосинтеза синезеленой водоросли. Некоторые виды Rhizosolenia, содержащие в цитоплазме нити анабены, в отличие от Geosiphon не утрачивают собственных пластид[27]. ____________________________
[27] Описанные случаи внутриклеточного симбиоза у водорослей и так
называемый лишайниковый (внеклеточный) симбиоз представляют большой
теоретический интерес. Некоторые из подобных примеров в свое время послужили
стимулом для зарождения теории симбиогенеза.
Экологические группировки высших водных растений. Высшие водные оксигенные фототрофы, или высшая водная растительность всегда интересовала не только натуралистов и исследователей, но и простых крестьян. И для тех, и для других была важна продуктивность и сельскохозяйственная значимость этих растений. С эволюционной точки зрения высшие водные растения можно рассматривать как группу преимущественно травянистых наземных растений, вторично перешедших к жизни в водных экосистемах. У одних из них весь жизненный цикл проходит только в водной среде, у других генеративные побеги могут возвышаться над поверхностью воды. У третьих в процессе онтогенеза возможны изменения в контактах с водной и воздушной средами.

|
|||
|
| ||