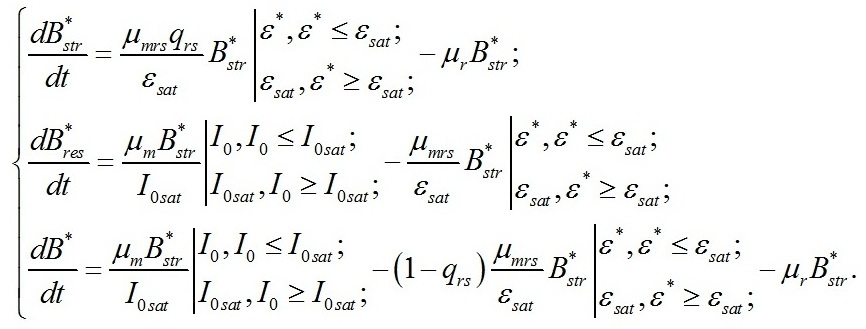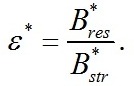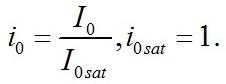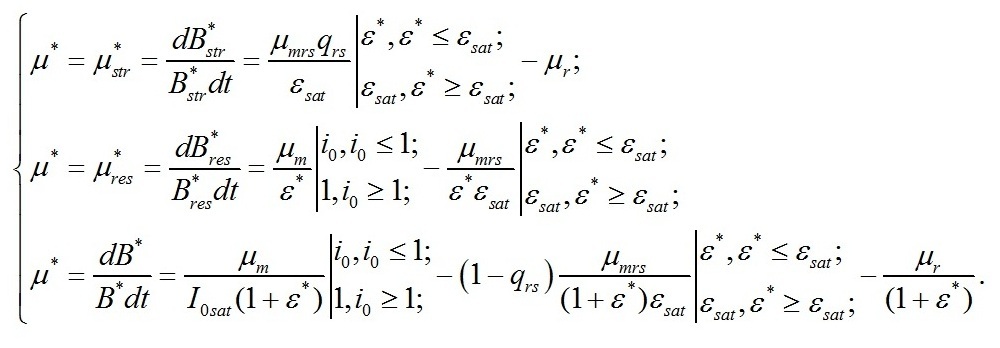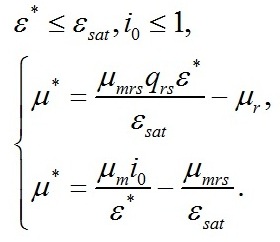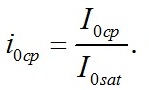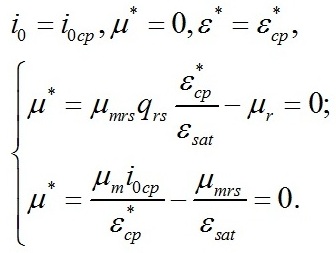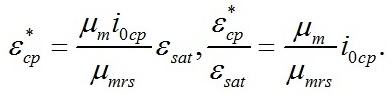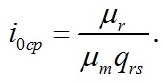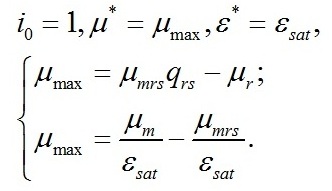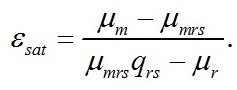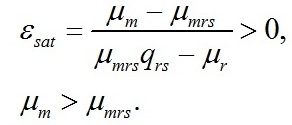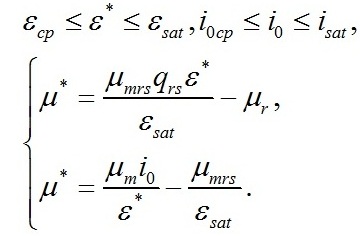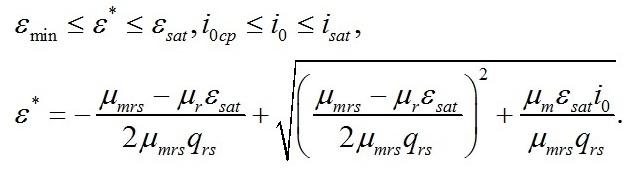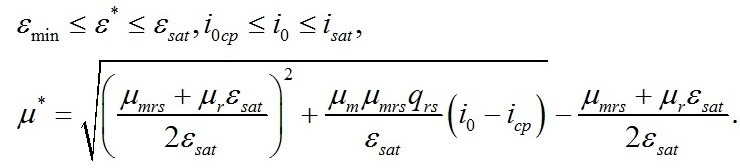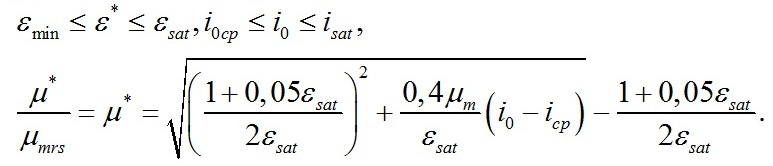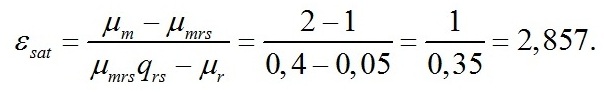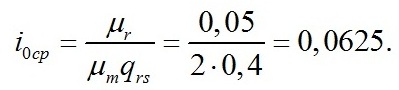|
|
|
|
по Материалам Международной конференции «Экологическая физиология водных фототрофов: распространение, запасы, химический состав и использование» VII Сабининские чтения 1 ноября - 25 декабря 2018 г. Влияние света на рост микроводорослей в непрерывной культуре невысокой плотности
Тренкеншу Р.П. Rudolf P. Trenkenshu
Институт морских биологических исследований им. А.О.Ковалевского РАН
УДК 581.1:57.032:57.033
Предложена математическая модель зависимости удельной скорости роста микроводорослей от интенсивности света для непрерывной культуры невысокой оптической плотности. В основе механизма светолимитированного роста лежит положение о разделении световой и темновой стадии фотобиосинтеза. Продуктом первой стадии являются углеводы, которые используются в темновой стадии в качестве источника энергии и углеродных скелетов для синтеза всех структурных компонентов клеток. Скорости процессов описываются линейными сплайнами. Получены уравнения, описывающие зависимость удельной скорости роста от интенсивности действующего света. Ключевые слова: микроводоросли; культивирование; скорость фотосинтеза; удельная скорость роста; биохимический состав.
Введение Главным фактором, определяющим фотосинтез микроводорослей при фотоавтотрофном росте в культуре, является свет. Для количественной оценки влияния света на рост и фотосинтез микроводорослей предложено много моделей, в которых используются те или иные параметры культур. Предлагаемая работа не ставит своей целью анализ известных моделей и получаемых при этом уравнений. Такой анализ можно найти в обзорах (Jassby, Platt, 1976; Белянин, 1984; Лелеков, 2009). Рассматривая влияние света на синтез биомассы микроводорослей в фотоавтотрофной культуре необходимо учитывать последовательность световых и темновых реакций, при которых ассимилируется минеральный углерод и за счет световой энергии в цикле Кальвина преобразуется в полисахариды. В свою очередь, эти первичные углеводы служат источником энергии и углеродных скелетов для всех последующих процессов биосинтеза клеточных структур (Физиология растений, 2005). Ясно, что такая последовательность процессов преобразования минеральных веществ в структурные компоненты клеток указывает на значительные биохимические изменения в составе биомассы. Огромное количество актов биохимических преобразований предполагает их учет для построения полной динамической модели этих процессов, что приведет к огромному количеству уравнений и бессмысленности моделирования в целом. В то же время для корректного описания биохимических трансформаций при стационарном росте достаточно учесть ключевые параметры, определяющие конечные скорости процессов. Большинство исследователей склоняются к тому, что самой медленной стадией фотосинтеза является активность первого и главного компонента цикла Кальвина – рибулезобифосфаткарбоксилазы – Rubisco (Portis, Parry, 2007). Кроме того, в клетках микроводорослей можно выделить структурную часть биомассы, которая участвует в процессах биотрансформаций в качестве катализаторов. Структурная часть биомассы также включает и Rubisco. Это позволяет принять пропорциональность между концентрациями ферментов и структурной биомассой и свести к минимуму количество уравнений. В предлагаемой работе рассматривается светозависимый фотоавтотрофный рост микроводорослей в динамически равновесной (непрерывной) культуре невысокой плотности. Выбор невысоких плотностей культур обусловлен тем, что позволяет избежать влияния «самозатенения» клеток, а используемый общий подход к моделированию роста микроводорослей в культуре сформулирован ранее в работе (Лелеков, Тренкеншу, 2018).
Система уравнений В основу работы положена система уравнений, предложенная ранее (Тренкеншу, 2016, 2017):
Надстрочные символы (*) означают, что рассматривается непрерывная динамически равновесная культура микроводорослей. В уравнениях приняты следующие обозначения. Биомасса микроводорослей представлена как сумма структурных и резервных (ресурсных) форм:
Соотношение этих форм:
Кинетические коэффициенты: Световые условия представлены в виде: Световые условия можно представить в безразмерном виде:
εsat – соотношение ресурсные/структурные формы биомассы при световом насыщении.
Учитывая, что в непрерывной культуре все удельные скорости равны и равны удельной скорости роста (или протока), получим:
Удельная скорость роста при световом лимитировании Для данных условий система редуцируется до вида:
Определим границы применимости для полученной системы уравнений. Непрерывный рост микроводорослей в фотоавтотрофной культуре возможен только при интенсивности света выше определенной величины, называемой компенсационным пунктом фотосинтеза (I0cp):
В точке светового компенсационного пункта скорость роста будет равной нулю:
Из уравнений находим величину соотношения ресурсные/структурные формы биомассы в точке, соответствующей световому компенсационному пункту при фотоавтотрофном росте микроводорослей. Из первого уравнения системы находим соотношение ресурсные/структурные формы биомассы в световой компенсационной точке. В этом случае величина соотношения будет минимальной для непрерывной культуры:
Эти же величины можно выразить через световой компенсационный пункт:
Второе уравнение также позволяет выразить величину светового компенсационного пункта через кинетические коэффициенты:
Найденная величина интенсивности света является нижней границей применения системы уравнений. Верхней границей служит насыщающая интенсивность света (I0sat), при которой цикл Кальвина (точнее, активность Rubisco) работает с максимальной скоростью. Удельная скорость роста микроводорослей при этом достигает максимального значения:
Полученная система уравнений позволяет выразить величину соотношения ресурсные/структурные формы биомассы при достижении максимальной скорости роста через кинетические коэффициенты:
Заметим, что последняя формула накладывает ограничения на соотношение кинетических коэффициентов, т.к. по смыслу соотношение может быть только положительным:
Найденные величины представляет собой верхнюю границу применимости системы уравнений:
Зависимость соотношения ресурсных и структурных макромолекулярных форм в биомассе микроводорослей от интенсивности света в пределах найденных границ будет описывается уравнением (Тренкеншу, 2017):
Подставляя это выражение в первое уравнение последней системы, и произведя некоторые преобразования, получим зависимость удельной скорости роста микроводорослей от интенсивности света в заданном диапазоне:
Графические представления В качестве примера рассмотрим, как зависит форма световой кривой при использовании кинетических коэффициентов, значения которых были приняты в работе (Тренкеншу, 2017). Учитывая, что максимальная скорость роста и фотосинтеза более, чем на порядок выше удельной скорости поддержания, примем, что их отношение отличается в 20 раз. Экономический коэффициент qrs примем равным 40%. Приведя удельную скорость роста к безразмерному виду по отношению к максимальной скорости расхода углеводов (приняв последнюю за единицу), получим новую запись уравнения для скорости:
Отношение максимальной удельной скорости фотосинтеза к максимальной скорости расхода углеводов примем равным 2. Такое значение удовлетворяет условию положительных значений εsat:
Находим компенсационный пункт фотосинтеза:
Максимальная удельная скорость роста:
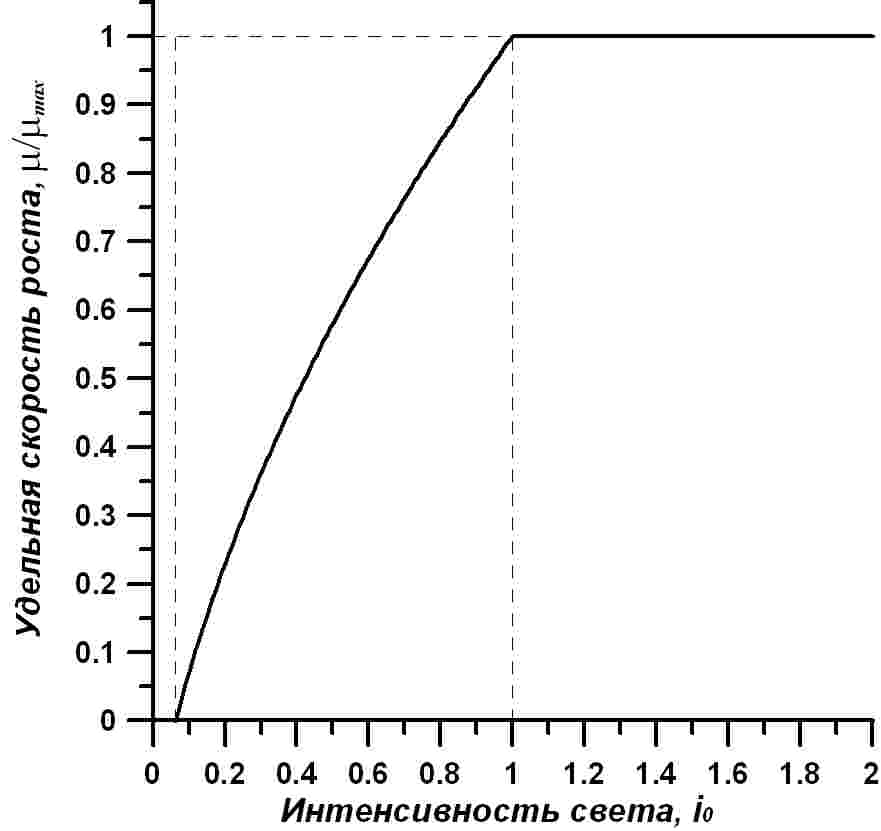
Расчет по уравнению зависимости удельной скорости роста от интенсивности света, выраженный в нормированном относительно максимального значения виде, показан на рисунке.
Рис. 1. Зависимость нормированных значений удельной скорости роста непрерывной культуры микроводорослей от интенсивности света Fig. 1. Dependence of normalized values of specific growth rate of continuous culture of microalgae on the light intensity
Заключение Используя общепринятое положение о разделении световой и темновой стадий фотобиосинтеза, получена математическая модель зависимости удельной скорости роста микроводорослей от интенсивности света для непрерывной культуры невысокой оптической плотности. Форма световой кривой в области светолимитированного роста в целом соответствует известным экспериментальным данным. В то же время, при насыщающих интенсивностях света уравнение не показывает наличие фотоингибирования, характерного для роста микроводорослей в непрерывной культуре. Такого рода вопросы требуют дальнейшего изучения.
Автор заявляет об отсутствии конфликта интересов, требующего раскрытия в данной статье. Работа выполнена в рамках гос. задания № 1001-2014-0017 «Разработка научных основ решения гидробиологических и биотехнологических проблем интегрированного управления прибрежными зонами»
Список литературы
Статья поступила в редакцию 21.12.2018
Об авторе Тренкеншу Рудольф Павлович – Trenkenshu Rudolf P. кандидат биологических наук r.trenkenshu@rambler.ru Корреспондентский адрес: Россия, 299011, Севастополь, пр. Нахимова, 2, ФГБУН ИМБИ; тел. (869)-2550795.
ССЫЛКА НА СТАТЬЮ: Тренкеншу Р.П. Влияние света на рост микроводорослей в непрерывной культуре невысокой плотности // Вопросы современной альгологии. 2019. № 1 (19). С. 1–7. URL: http://algology.ru/1395 DOI - https://doi.org/10.33624/2311-0147-2019-1(19)-1-7 Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно. При перепечатке ссылка на сайт обязательна
Influence of light on microalgae growth in continuous culture of low density Rudolf P. Trenkenshu Kovalevsky Institute of Marine Biological Research of RAS (Sevastopol, Russia) A mathematical model of the dependence of the specific growth rate of microalgae on the light intensity for a continuous culture of low optical density is proposed. In the basis of the mechanism of light-limited growth is the thesis on photosynthesis having light and dark phases. The product of the first phase is carbohydrates, which are used in the dark phase as a source of energy and carbon skeletons for the synthesis of all structural components of cells .Rates of the processes are described by linear splines. The equations describing the dependence of the specific growth rate on the intensity of the acting light are obtained. Key words: microalgae; cultivation; photosynthesis rate; specific growth rate; biochemical composition.
Author Trenkenshu Rudolf P. ORCID - https://orcid.org/0000-0003-3727-303X Kovalevsky Institute of Marine Biological Research of RAS, Sevastopol, Russia r.trenkenshu@rambler.ru
Trenkenshu R.P. Influence of light on microalgae growth in continuous culture of low density. Voprosy sovremennoi algologii (Issues of modern algology). 2019. № 1 (19). P. 1–7. URL: http://algology.ru/1395 DOI - https://doi.org/10.33624/2311-0147-2019-1(19)-1-7
References
Dear colleagues! If you want to receive the version of the article in PDF format, write to the editor,please and we send it to you with pleasure for free. When reprinting a link to the site is required
К разделу ОБЗОРЫ, СТАТЬИ И КРАТКИЕ СООБЩЕНИЯ

|
|||
|
| ||