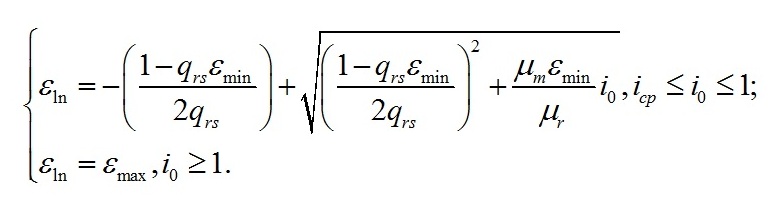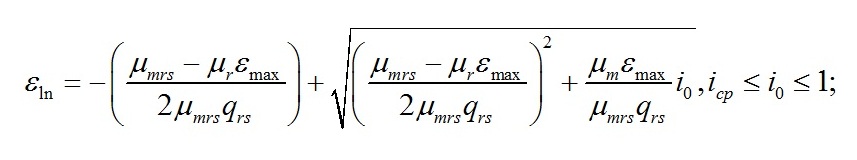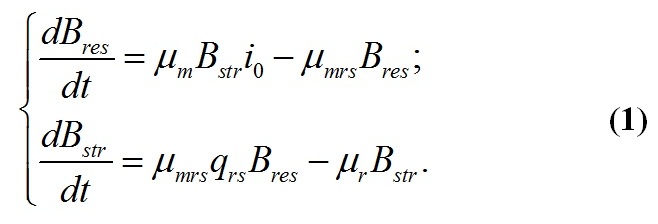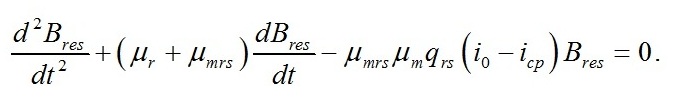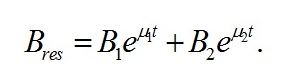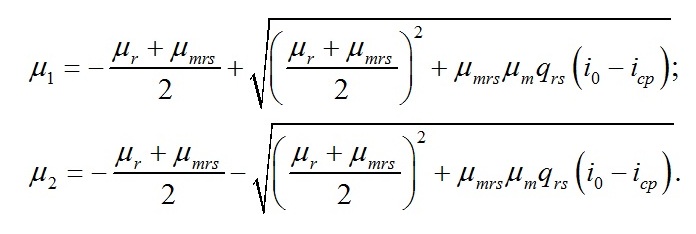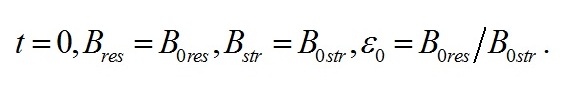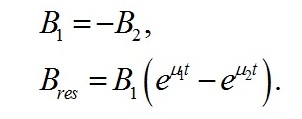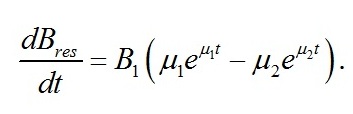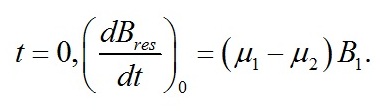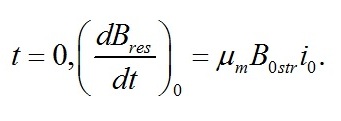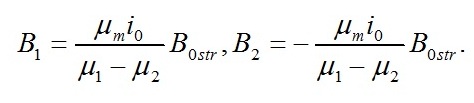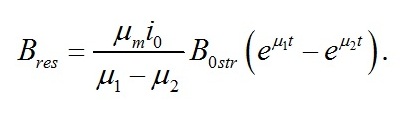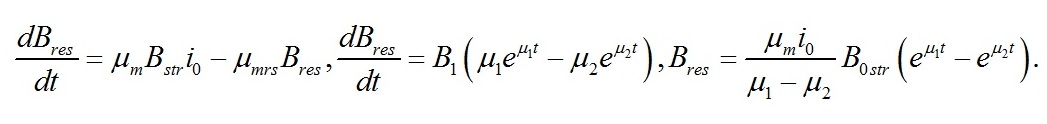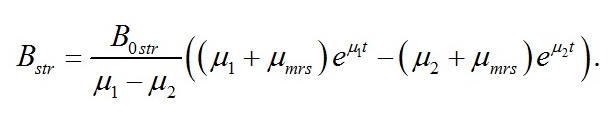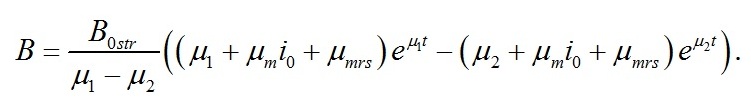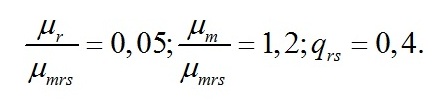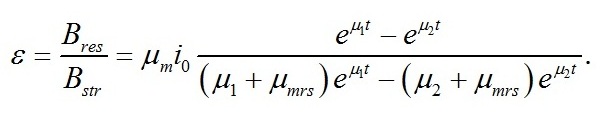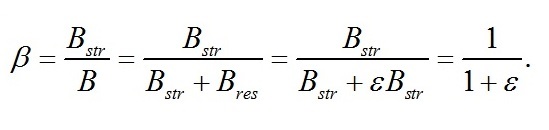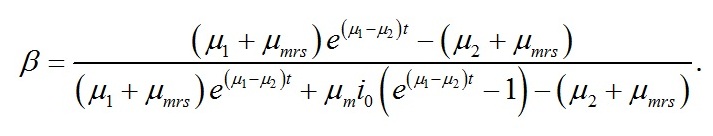|
|
|
Рост микроводорослей при переходе от темноты к постоянному освещению
Тренкеншу Р.П. Rudolf P. Trenkenshu
Институт морских биологических исследований им. А.О. Ковалевского (Севастополь, Россия)
УДК 581.1:57.032:57.033
В статье рассматривается биохимическая адаптация культуры микроводорослей к постоянному освещению после длительного содержания клеток в темноте. Начало лаг-фазы охарактеризовано отсутствием резервных форм биомассы, т.е. клеточное вещество представлено только структурными компонентами клеток. Предложены уравнения, описывающие изменения во времени концентраций резервных и структурных форм, а также биомассы (как их суммы). Показано, что их соотношения изменяются вплоть до перехода культуры к экспоненциальному росту, характеризующемуся постоянством соотношений. Приведены графические иллюстрации моделей для случая перехода от темноты к насыщающей интенсивности действующего света. Ключевые слова: микроводоросли; биохимический состав; скрытая фаза роста; экспоненциальный рост; моделирование.
Введение Наиболее распространенным способом исследования роста микроводорослей в зависимости от различных факторов внешней среды является метод периодической (накопительной) культуры. Накопительная культура характеризуется последовательностью фаз роста, причем первая фаза – скрытая (лаг-период) – наименее изученная. В этой фазе роста клетки адаптируются к новым заданным условиям среды (Anning et al., 2000). После адаптации, при неизменной интенсивности света, культура переходит к экспоненциальной или линейной фазе роста, это зависит от плотности инокулята и интенсивности внешнего света. Если плотность культуры невелика, а концентрация элементов питания достаточна для продолжения интенсивного роста, то культура перейдет в экспоненциальную фазу, которая характеризуется постоянством удельной скорости роста, определяемой только световыми условиями. Постоянство удельной скорости роста требует неизменности соотношения всех без исключения химических составляющих биомассы (Тренкеншу, 2009). Это означает, что до перехода культуры в экспоненциальную фазу роста в клетках должен сформироваться определенный биохимический состав, соответствующий заданным световым условиям. Такое формирование возможно только при различных удельных скоростях синтеза отдельных химических составляющих в биомассе микроводорослей в лаг-фазе роста культуры. Особенно заметные изменения биохимического состава культуры в лаг-фазе происходят при использовании инокулята, относительно долгое время находящегося в темноте или при интенсивностях света ниже компенсационного пункта фотосинтеза. Это связано с «темновой потерей биомассы» в фотоавтотрофной культуре микроводорослей (Eppley, Sharp, 1975, Edmundson, Huesemann. 2015). Показано, что в клетках планктонных микроводорослей ночью происходит окисление углеводов (Eppley, Sharp, 1975), а в культуре микроводорослей наблюдается темновое дыхание (Grobbelaar, Soeder, 1985). Причем скорости процессов относительно высокие (Eppley, Sharp, 1975; Grobbelaar, Soeder, 1985). Вместе с этим во время нахождения клеток в темноте, кроме резкого снижения содержания углеводов в культуре микроводорослей наблюдается накопление белков и некоторое снижение количества липидов (Hu et al., 1996, Ogbonna, Tanaka, 1996). Иными словами, в темноте происходят значительные биохимические изменения состава биомассы. Количественно эти изменения могут быть выражены через соотношение резервных и структурных форм биомассы (Тренкеншу, 2016) . Как показано в работе (Тренкеншу, 2009), для подтверждения экспоненциального роста культуры микроводорослей на некотором промежутке времени достаточно постоянства соотношения двух биохимических характеристик культуры на этом промежутке. Такое свойство экспоненциального роста позволяет выделить из состава биомассы любые макромолекулярные формы биомассы и изучить зависимость их соотношения от интенсивности действующего света. Например, содержания белка или пигментов в биомассе клеток. Эти относительные величины зависят от световых условий и могут быть описаны количественно (Anning et al., 2000, Тренкеншу, 2017), т.е. появляется возможность оценки биохимического состава культуры, к которому приведет внешний свет при переходе к экспоненциальному росту. А зная биохимический состав инокулята, можно определить форму зависимости этого перехода в лаг-фазе роста культуры. В данной работе представлены результаты моделирования биохимической адаптации клеток во время перехода культуры от темновых условий к экспоненциальному росту на свету.
Макромолекулярный состав клеток при экспоненциальном росте Переход в экспоненциальную фазу роста будет происходить до тех пор, пока биомасса не достигнет соотношений состава, определяемого световыми условиями. В качестве основы для моделирования в работе (Тренкеншу, 2016) были выбраны две формы состава биомассы: структурная (Bstr) и резервная (Bres), сумма которых представляет собой биомассу (B). Этот выбор не случаен, т.к. согласно общепринятым представлениям о фотосинтезе, ассимиляция углерода зависит от света и накапливается в клетках в виде резервных (=ресурсных) полисахаридов, которые затем трансформируются в структурные компоненты клеток (Мокроносов, 2006; Физиология растений, 2007). Исходя из таких представлений, биомассу и её макромолекулярные формы удобнее всего выражать в виде углеродных единиц. В работе (Тренкеншу, 2017) показано, что при экспоненциальном росте культуры соотношение резервных и структурных форм биомассы (εln=lnBres/ lnBstr) зависит от интенсивности света (i0):
Коэффициент µm представляет собой активность рибулозобисфосфат-карбоксилазы/оксигеназы (Rubisco) и включает долю этого фермента в структурной биомассе и экономический коэффициент преобразования световой энергии в химическую энергию продуктов цикла Кальвина. Активность фермента (или транспортной системы) в реакциях синтеза структурных компонентов клетки из ресурсной формы выражена коэффициентом µmrs. Коэффициент qrs – экономический коэффициент трансформации в реакциях синтеза структурных компонентов клетки из ресурсной формы. µr – удельная скорость поддержания структуры клетки (Перт, 1978). Система уравнений включает как кинетические коэффициенты, так и видоспецифические параметры микроводорослей, которые могут быть выражены через эти коэффициенты и внешние условия. К видоспецифическим параметрам относятся величины максимального (εmax) и минимального (εmin) соотношения ресурсной и структурной форм биомассы при экспоненциальном росте. Отношение этих предельных характеристик определяется совокупностью кинетических коэффициентов:
Используя это соотношение первое уравнение системы можно записать в другой форме:
Единственным переменным внешним параметром в уравнениях является свет. Остальные условия считаются неизменными и не влияющими на рост микроводорослей. Интенсивность света (i0) выражена в виде безразмерных единиц относительно насыщающей величины. Максимальное соотношение (εmax) наблюдается при интенсивности света выше или равной насыщающему значению (i0 ≥ 1). Минимальное соотношение (εmin) характеризует предельно малое значение этой величины в точке светового компенсационного пункта (i0 = iср) фотосинтеза, который может быть выражен через кинетические коэффициенты:
Таким образом, сформулированы условия, при которых возможен экспоненциальный рост. Для старта экспоненциальной фазы в накопительной культуре микроводорослей необходимо, чтобы к этому моменту времени макромолекулярный состав биомассы пришел к необходимым пропорциям, соответствующим интенсивности действующего света.
Динамика макромолекулярного состава клеток Приведенная выше система уравнений является стационарным решением системы уравнений, описывающей фотосинтез ресурсных форм и их переход в структурные формы биомассы (Тренкеншу, 2016, 2017), которая с несущественным упрощением может быть записана в виде:
Решение такого типа систем уравнений сводится к преобразованию системы в дифференциальное уравнение второго порядка через выражение одной переменной в другую путем подстановок.
Динамика ресурсных форм биомассы Для ресурсной формы биомассы уравнение второго порядка имеет вид:
В случае интенсивностей света, превышающих световой компенсационный пункт, решение последнего уравнения имеет вид:
В уравнении приняты обозначения:
Как и следовало ожидать, коэффициенты при показателе экспонент представляют собой удельные скорости изменения ресурсных форм биомассы, выражаются через кинетические коэффициенты и зависят от интенсивности света, к которой адаптируются микроводоросли. Для определения констант интегрирования (В1 и В2) воспользуемся данными о начальных условиях. В начальный момент времени (t=0) микроводоросли имеют начальный макромолекулярный состав биомассы:
Подставляя начальное значение ресурсной формы биомассы в интегральное решение, получим его связь с постоянными интегрирования:
В данной работе рассматривается случай использования инокулята после относительно длительного содержания микроводорослей в темноте или при низких интенсивностях освещения. Т.е. начальное значение резервных веществ в клетке близко к нулю:
Имеем:
Из последнего уравнения находим скорость изменения ресурсной формы биомассы:
В начальный момент времени:
Эта же скорость из первого уравнения системы (1):
Сравнивая два последних уравнения, находим константы интегрирования:
Окончательно имеем:
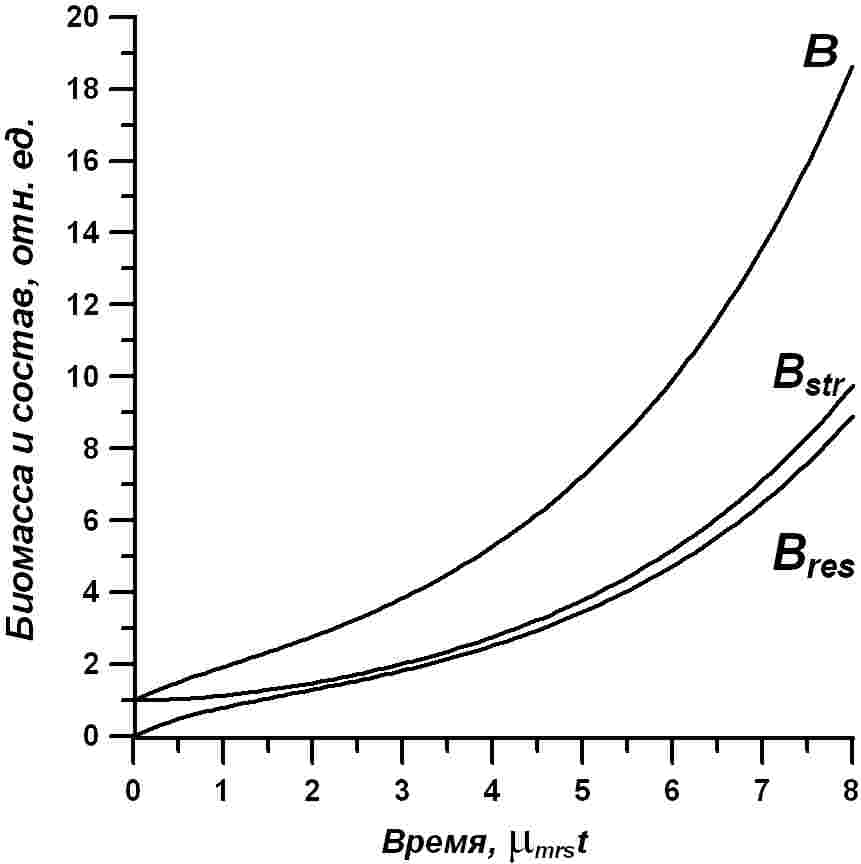
На рис. 1 показано накопление резервных форм биомассы в культуре микроводорослей при переходе от темноты к постоянному освещению.
Динамика структурных форм биомассы Накопление в культуре микроводорослей структурных компонентов клеток может быть найдено из первого уравнения системы (1) путем подстановки уравнения для скорости изменения ресурсных форм:
В итоге получим:
Графически полученная зависимость представлена также на рис. 1.
Динамика плотности культуры Концентрация биомассы в культуре представляет собой сумму концентраций резервных и структурных биохимических составляющих:
Подставляя полученные уравнения для разных форм биомассы в последнюю формулу, после преобразований получим уравнение динамики плотности культуры:
Расчет по этому уравнению также представлен на рис. 1. В качестве примеров все графические иллюстрации даны для случая, когда культура микроводорослей после содержания в темноте переводится на постоянный свет насыщающей интенсивности (i0=1). Для расчета всех кривых использовались коэффициенты из работы (Тренкеншу, 2017):
Расчет концентрации биомассы и её форм нормирован относительно начальной концентрации структурных форм (B0str). Время также приведено в нормированном относительно (1/μmrs) виде.
Рис. 1. Динамика накопления биомассы (B) и её структурных (Bstr) и резервных (Bres) составляющих в культуре микроводорослей. Пояснения в тексте Fig. 1. Dynamics of biomass accumulation (B) and its structure (Bstr) and reserve (Bres) components in microalgae culture. Explanations in the text
Соотношение ресурсные/структурные формы Важной видоспецифической характеристикой, определяющей скорости роста микроводорослей, является соотношение резервных и структурных составляющих биомассы. Это связано с тем, что резервные формы в химическом смысле представляют собой субстрат, из которого в результате ферментативных реакций продуцируется структурное вещество клеток. А мультиферментные комплексы (включая транспортные системы клетки) являются структурными компонентами клеток. Из полученных уравнений находим формулу для описания изменения соотношения ресурсные/структурные формы биомассы во время адаптации клеток (лаг-период или скрытая фаза) и при переходе к экспоненциальной фазе роста:
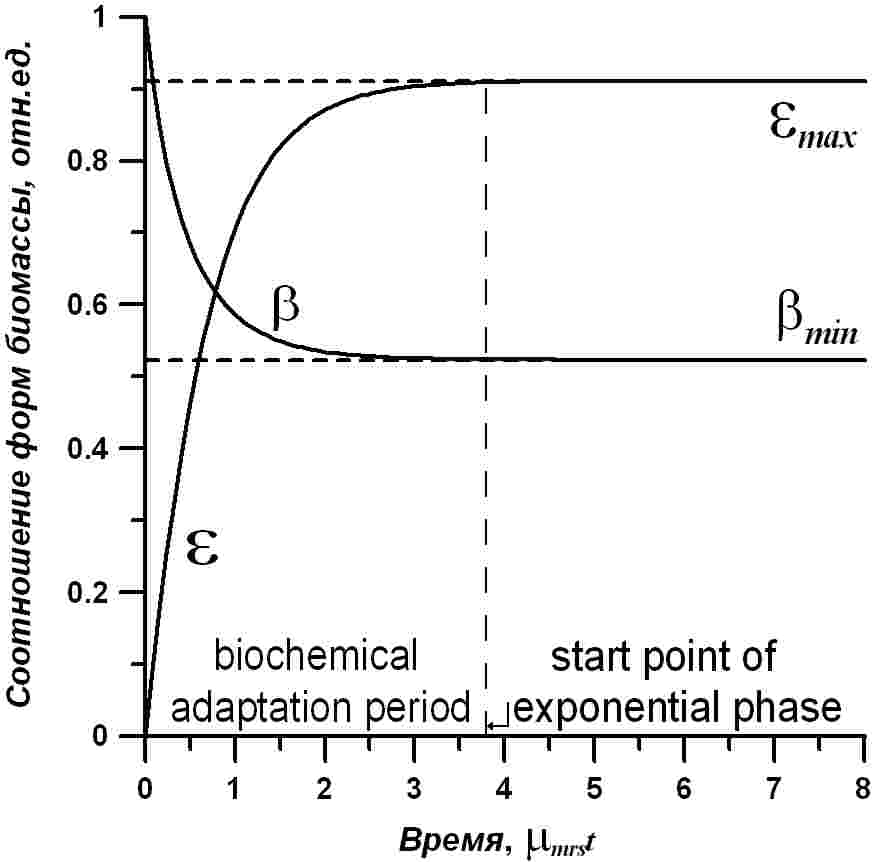
Графическая иллюстрация расчета динамики соотношения ресурсные/структурные формы биомассы показана на рис. 2. Видно, что это соотношение изменяется со временем от нулевых значений (темнота) до стационарных величин, определяемых световыми условиями. В данном примере стационарное значение соотношения указано как максимальное, т.к. расчет произведен для насыщающей интенсивности света (i0=1). Задавая различные величины действующего света в полученном уравнении можно получить семейство кривых, описывающих биохимические изменения в лаг-фазе роста культуры микроводорослей, а также оценить время окончания адаптационного периода и перехода к экспоненциальному росту.
Рис. 2. Динамика изменения соотношения резервных и структурных форм (ε) и доля структурных составляющих в биомассе микроводорослей (β). Выделена область начала и конца адаптации (лаг-фаза роста) и точка перехода в экспоненциальную фазу. Пояснения в тексте Fig. 1. Dynamics of changes in the ratio of reserve and structure forms (ε) and the share of structural components in microalgae biomass (β). The region of the beginning and the end of adaptation (lag-phase of growth) and transition point to the exponential phase is shown. Explanations in the text
Содержание структурных форм в биомассе В практике изучения биохимического состава микроводорослей довольно часто пользуются величиной, характеризующей содержание белка в биомассе. Так как белки относятся к структурным компонентам клеток, можно принять, что доля белков в структуре клеток является постоянной величиной. В этом случае характер динамической кривой изменения содержания белка в биомассе будет аналогичен динамической кривой изменения структурных форм в биомассе (β) со временем:
Такая простая форма перехода от соотношения ресурсные/структурные составляющие приводит к виду уравнение для содержания структурных форм в биомассе:
Уравнение справедливо для случая помещения микроводорослей на постоянный свет после содержания в темноте. На рис. 2 показана кривая изменения содержания структурных форм в биомассе при переходе от темноты к насыщающему свету. Видно, что за время адаптации содержание структурных составляющих в клетках изменяется от стопроцентного количества до стабильных минимальных значений после начала экспоненциального роста. Уровень этого минимума будет зависеть от интенсивности света. На рис. 2 также показана длительность лаг-фазы и точка начала экспоненциального роста.
Заключение В работе рассмотрен частный случай биохимической адаптации микроводорослей в начальной фазе роста при их выращивании в культуре. Этот участок накопительной (периодической) кривой роста микроводорослей носит название лаг-фаза и практически не изучен, хотя большинство исследовательских работ с микроводорослями проведено именно с применением такого метода культивирования. В настоящей работе показано, что в этом периоде роста происходят значительные изменения биохимического состава, а соотношения составляющих биомассы могут изменяться во много раз. При проведении экспериментальных исследований методом накопительных культур необходимо учитывать эти различия.
Работа выполнена в рамках гос. задания № АААА-А18-118021350003-6. «Разработка научных основ решения гидробиологических и биотехнологических проблем интегрированного управления прибрежными зонами».
Список литературы
Статья поступила в редакцию 24.10.2018
Microalgae growth at transition from darkness to constant lighting Rudolf P. Trenkenshu The article focuses on biochemical adaptation of microalgae culture to constant lighting after being kept in the dark conditions for a long time. The beginning of the lag phase is characterized by the absence of reserve forms in the biomass, i.e. it is presented only by structural components of cells. Equations describing changes in time of the concentrations of reserve and structure forms, as well as biomass (as their sum) are proposed. It is shown that their ratio changes up to the point when the culture starts its exponential growth, characterized by constant ratio. Graphic illustrations of the models for the case of transition from darkness to saturating intensity of the incident light are given. Key words: microalgae; biochemical composition; latent growth phase; exponential growth; modeling.
Об авторе Тренкеншу Рудольф Павлович – Trenkenshu Rudolf P. кандидат биологических наук r.trenkenshu@rambler.ru Корреспондентский адрес: Россия, 299011, Севастополь, пр. Нахимова, 2, ФГБУН ИМБИ; тел. (869)-2550795.
ССЫЛКА НА СТАТЬЮ: Тренкеншу Р.П. Рост микроводорослей при переходе от темноты к постоянному освещению // Вопросы современной альгологии. 2018. № 2 (17). URL: http://algology.ru/1350 Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно.
При перепечатке ссылка на сайт обязательна
На ГЛАВНУЮ
|
|||
|
| ||