|
|
|
Сообщества бурых водорослей рода Cystoseira северо-восточной части Черного моря
Афанасьев Д.Ф.1,2, Камнев А.Н.3, Стуколова И.В.3 Dmitry F. Afanasyev1,2, Alexander N. Kamnev3, Irina V. Stukolova3
1Азовский научно-исследовательский институт рыбного хозяйства (Ростов-на-Дону)
581.9(262.5)
Исследованы сообщества бурых водорослей рода Cystoseira на шельфе северо-восточного побережья Черного моря. Выявлена структура сообществ C. crinita на глубинах 2, 5 и 10 м. Установлено, что сезонность проявляется в основном в изменении биомассы вида-доминанта – C. crinita – которая имеет один сезонный максимум (май-июнь) и минимум (февраль). Сезонная динамика биомассы Cystoseira barbata аналогична динамике C. crinita, однако проявляется только на глубине 2 м. Сезонная динамика сообществ на глубине 10 м для большинства исследованных параметров не выявлена. Ключевые слова: Черное море; фитобентос; Cystoseira; структура; биомасса; общее проективное покрытие.
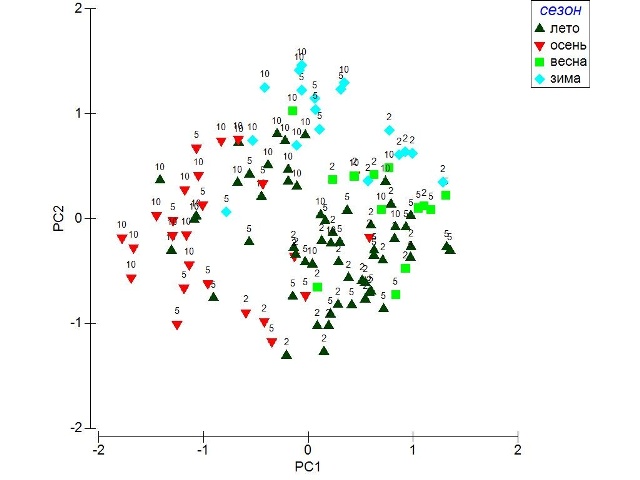
Сообщества с доминированием морских водорослей рода Cystoseira порядка Fucales являются одним из основных элементов растительности сублиторальной зоны Средиземного, Мраморного и Черного морей (Giaccone, 1973; Giaccone et al., 1993, 1994; Калугина-Гутник, 1975; Камнев, 1989; Ballesteros, 1990a, 1990b, 1992; Montesanto, Panayotidis, 2001; Panayotidis et al. 2007). Согласно последним данным ДНК-секвенирования, распространение рода Cystoseira ограничено южными морями Европы (Draisma et al. 2010), что делает подводные заросли этих бурых водорослей уникальными, учитывая их наиболее высокое биологическое разнообразие в системе всей макроскопической растительности указанных акваторий. Растительность Черного моря является одной из наименее изученных среди других внутренних морей Атлантического бассейна. Такие особенности Черного моря, как относительно низкая соленость (в среднем у северо-восточного шельфа – 18‰), низкие температуры воды и широкий диапазон их колебаний на мелководных участках, изолированность от других морей и океанов, обусловливают довольно низкое биологическое разнообразие макрофлоры. Из почти 50 видов рода Cystoseira, доминирующих на шельфе Средиземного моря и прилегающих районах Атлантического Океана, в Черном море встречаются 5, а в его северо-восточной части – только 2 вида (Ribera et al., 1992; Мильчакова, 2002а, 2002b). Тем не менее, эти 2 вида – Cystoseira crinita и Cystoseira barbata – играют основную роль в формировании донных растительных сообществ в Черном море, а черноморские сообщества с доминированием C. crinita являются наиболее богатыми во флористическом отношении (Калугина-Гутник, 1975). Если в Средиземном море разные виды цистозир занимают разные местообитания в зависимости от глубины, угла экспозиции по отношению к волновой активности и других экологических факторов (Giaccone, 1971, Ballesteros, 1992, Sales, Ballesteros, 2009), то в Черном море глубины от 0,5 до 15-20 м заняты только сообществами C. crinita и C. barbata (Калугина-Гутник, 1975; Громов, 1998; Афанасьев, Корпакова, 2008). Вопрос о синтаксономическом положении этих сообществ до сих пор остается открытым, так как, несмотря на сходство черноморских сообществ цистозир со средиземноморскими ассоциациями Cystoseiretum crinitae Molinier 1958 и Cystoseiretum barbatae Pignatti 1962 (Molinier, 1958; Pignatti, 1962; Giaccone, Pignatti, 1967), их пространственное распределение, экологические особенности и состав диагностических видов не всегда соответствует таковому в Средиземном море (Афанасьев и др., 2012). В первую очередь это касается более широкой экологической амплитуды этих ассоциаций в Черном море и значительно большего проективного покрытия C. barbata в черноморских сообществах C. crinita. Во избежание путаницы в описании сообществ, которые с большой долей вероятности являются разными синтаксонами, в настоящей статье авторы рассматривают только те сообщества, в которых проективное покрытие C. crinita значительно преобладает над проективным покрытием C. barbata. Сезонная динамика сообществ C. crinita исследовалась лишь в западной части Средиземного моря, на побережье Сицилии (Pizzuto, 1999), Каталонии (Ballesteros, 1992) и на Балеарских островах (Sales, Ballesteros, 2012). Эти исследования проводились в зоне инфралиторали, на глубинах 0-1 м, так как глубже средиземноморские сообщества C. crinita сменяются на сообщества с доминированием других видов рода Cystoseira. Для Черного моря вопрос о комплексном исследовании сезонной динамики сообществ C. crinita на всех глубинах их произрастания не ставился, так как эти сообщества традиционно разделялись на несколько ассоциаций, объем которых трактовался авторами по-разному. Преимущественно исследовалась сезонная динамика видового состава сообществ (Калугина-Гутник, 1975) и биомасса отдельных фитоценозов с доминированием C. crinita, выделяемых в качестве отдельных ассоциаций и не соответствующих объему сообществ с доминированием C. crinita (Громов, 1998). Важность исследований прибрежных донных фитоценозов и ассоциированных с ними экологических комплексов обусловлена их высокой продуктивностью (благодаря обилию биогенов, высокой температуре, освещенности, разнообразию биотопов), высоким уровнем α-, β- и γ-разнообразия (как инвентаризационного, так и дифференцирующего), уникальностью, высокой (рыбо)хозяйственной и биологической значимостью (места нереста горбылей, саргана, бычков, атерины, нагула барабули, камбал, хамсы, сельди, луфаря, кефалей), высокой степенью изменчивости и уязвимости (риски острого и хронического загрязнения, эвтрофикации, уничтожения). Цель настоящего исследования – изучить сезонную динамику видового состава и структуры всех сообществ с доминированием C. crinita относительно чистых районов северо-восточной части Черного моря, рассматриваемых авторами в качестве единого синтаксона. Материалы и методы Для исследований сезонных вариаций видового состава и сырой биомассы сообществ C. crinita был выбран полуостров Абрау северо-восточной части Черного моря, донная флора и растительность которого отличается максимальным разнообразием и наибольшей глубиной проникновения. Исследования проводили на трех однородных по экологическим условиям разрезах: у пос. Южная Озереевка, пос. Лиманчик и пос. Сукко – м. Большой Утриш. Разрезы расположены у открытых берегов и ориентированы на юг и юго-запад. Исследовали глубины 2, 5 и 10 м на каждом разрезе. Грунт на разрезах скально-валунный, с микрорельефом 10-30 см, редко до 50 см. На каждой станции закладывали рамки размером 0,25 м2 (весной, летом и осенью) или 0,11 м2 (зимой). Из рамки отбирали все водоросли-макрофиты для определения видов и их биомассы в лаборатории. Для оценки сырой биомассы видов, водоросли промокали фильтровальной бумагой до воздушно-сухого состояния и взвешивали. Полученные результаты пересчитывали на 1,0 м2 (Воронов, 1973; Громов, 1973). Анализировалась биомасса водорослей «на заросли» (без учета общего проективного покрытия) и общая биомасса с учетом общего проективного покрытия (ОПП). Для мелких красных корковых видов водорослей и некоторых мелкоразмерных видов эпифитов биомасса не определялась. Пробы отбирали с использованием легководолазного снаряжения, как самостоятельно, так и с привлечением сертифицированных водолазов. В разные сезоны исследовали от 4 до 11 рамок, отобранных на каждой глубине. Исследования проводили в течение года в период с июня 2012 по май 2013 г.: 22-23 июня 2012 г., 5-6 и 20-21 августа 2012 г., 21-22 ноября 2012 г., 20-21 февраля 2013 г. и 26-27 мая 2013 г. Всего было проанализировано 108 проб. Для проверки гипотезы о влиянии глубины обитания водорослей и сезона на общую биомассу сообществ, биомассу видов C. crinita и C. barbata, биомассу диагностических видов сообществ использовали двухфакторный анализ вариант. Для проверки парных гипотез post hoc использовали тест Тьюки. Ординация была выполнена методом главных компонент на основании данных по флористическому составу сообществ (наличие/отсутствие вида). При создании матрицы сходства использовали индекс Брей–Кертиса. Результаты обработаны и визуализированы с использованием программных пакетов Statistica 6.0, Primer 5 и MS Excel. Результаты и обсуждение Всего в сообществах с доминированием C. crinita было определено 80 видов водорослей-макрофитов, в том числе 51 вид красных, 12 – бурых и 17 – зеленых. На глубине 2 м за весь период наблюдений было выявлено 56 видов, на глубине 5 м – 52 вида, на глубине 10 м – 51 вид. В течение всего года в верхнем ярусе доминировала C. crinita, на большинстве станций вместе с ней встречалась C. barbata. Во втором ярусе (или в качестве эпифита) преобладала бурая водоросль Cladostephus spongiosus. Круглый год и на всех исследованных глубинах в качестве эпифитов на цистозирах встречались Polysiphonia subulifera, Laurencia obtusa, Sphacelaria cirrosa, Chaetomorpha linum, Acrochaetium sp., Melobesia farinosa, Melobesia minutula, на валунах – Corallina elongata, Peyssonnelia rubra. Указанные виды (15% общего видового состава сообществ) могут считаться константными для сообществ с доминированием C. crinita северо-восточной части Черного моря. Остальные 85% видов встречались на отдельных глубинах или в отдельные сезоны года. Максимальное количество видов выявлено в мае (48 видов) и августе (45 видов), минимальное – в июне и ноябре (27 и 31 вид соответственно). В целом, в течение года в пробах сообществ C. crinita на разных глубинах количество видов менялось от 18 до 35. Ординация проб методом главных компонент объясняет менее 20% общей вариабельности флористического состава (рис.1).
Рис. 1. Ординационный анализ флористического состава сообществ цистозиры кустистой методом главных компонент, собранных на глубинах 2, 5 и 10 м в разные сезоны.
Общее проективное покрытие водорослей на глубине 2 и 5 м во все исследованные сезоны было близко к 90-100%. На глубине 10 м на дне появлялись участки не занятые растениями, что частично зависело от накопления рыхлых осадков. Расположение сообществ здесь носило мозаичный характер, а общее проективное покрытие фитобентоса менялось от 30 до 70%. Согласно результатам статистической обработки проб, оба исследованных комплекса факторов (сезон и глубина отбора проб) достоверно влияли на общую биомассу исследованных сообществ, биомассу C. crinita и C. barbata (как с учетом ОПП, так и биомассу «на заросли», т.е. без учета ОПП). С увеличением глубины, общая биомасса сообществ, биомасса C. crinita и C. barbata (как с учетом ОПП, так и «на заросли») достоверно снижаются во все сезоны года. Изменение общей биомассы сообществ C. crinita и биомассы C. crinita с учетом ОПП достоверно зависит от сезона только на глубине 2 и 5 м. Оба параметра имеют один сезонный минимум (февраль). На глубине 10 м влияния сезона отбора проб на общую биомассу сообщества и биомассу C. crinita не выявлено. Сезонные изменения биомассы C. barbata выявляются только на глубине 2 м, биомасса C. barbata на глубине 5 и 10 м достоверно не различаются. Характер вертикального распределения сообществ C. crinita в Черном море, таким образом, значительно отличаются от такового в Средиземном море. В последнем, сообщества C. crinita вегетируют в основном в зоне верхней инфралиторали, на глубине 0-1 м, а глубже замещаются сообществами с доминированием других видов этого рода (Giaccone et al., 1994; Sales, Ballesteros, 2012). Можно предположить, что в Черном море более широкий диапазон глубин, занятых сообществами C. crinita, является следствием перераспределения экологических ниш между C. crinita и C. barbata в условиях отсутствия других видов этого рода. Возможно, также, что экологические различия между C. crinita в Средиземном и Черном морях закреплены генетически. На глубинах 2 и 5 м общее проективное покрытие сообществ C. crinita в Черном море не меняется, однако к глубине 10 м оно снижается почти вдвое. Частично это можно объяснить накоплением рыхлых грунтов в понижениях между валунами. Однако на этой же глубине встречаются и валуны, не покрытые зарослями макрофитов, чего не наблюдается на меньших глубинах. Достоверное снижение с глубиной общей биомассы водорослей, биомассы C. crinita и C. barbata «на заросли» позволяет сделать вывод, что снижение биомассы происходит не только по причине уменьшения ОПП сообществ – в пятнах зарослей на глубине 10 м биомасса также была достоверно ниже, чем на глубине 2 и 5 м. Таким образом, изреживание зарослей сообществ C. crinita к глубине 10 м зависит от комплекса факторов – накопления рыхлых осадков и неидентифицированного фактора (или факторов), действие которого пропорционально глубине (возможно, освещенность, температура и т.п.). Большинство исследованных параметров фитоценозов имеют выраженную сезонную динамику лишь на глубинах 2 и 5 м. На глубине 10 м, очевидно, происходит стабилизация условий среды, сезонные флуктуации состава и структуры сообществ здесь либо слабо выражены, либо вовсе не наблюдаются. Исследованные сообщества C. crinita отличаются наибольшим количеством видов среди других сообществ макроводорослей северо-восточной части Черного моря. К исследованным нами сообществам наиболее близка ассоциация Cystoseira crinita + Cystoseira barbata – Cladostephus verticillatus – Corallina mediterranea, описанная А.А. Калугиной-Гутник (1975). В настоящее время, по сравнению с данными, полученными ею в 70-х гг. ХХ в., в составе сообществ C. crinita, произрастающих в относительно чистых районах моря, количество видов снизилось. Почти все другие черноморские сообщества Cystoseira (как правило, это мелководные (Cystoseira barbata + Cystoseira crinita – Dilophus fasciola) или глубоководные ((Cystoseira barbata) – Phyllophora crispa – Codium vermilara) сообщества с доминированием C. barbata) характеризуются более низким видовым разнообразием. Другой особенностью черноморских сообществ C. crinita является значительный вклад в общую биомассу C. barbata, доля которой составляет в среднем 22,1% от общей. Существенный вклад сопутствующих видов цистозир (например, Cystoseira compressa) в сообществах C. crinita показан и для некоторых районов Средиземного моря (Tsiamis et al. 2006; Panayotidis et al., 2007). Однако, по крайней мере, в Балеарском море, сопутствующие виды цистозир (Cystoseira spinosa, C. barbata) не достигают значительного развития в исследуемых сообществах (Ballesteros, 1992; Sales, Ballesteros, 2012). В целом, изменение общей биомассы сообществ C. crinita оказывается зависимым практически только от биомассы самой C. crinita, доля которой в среднем составляет 66,1% общей биомассы всего сообщества. Средняя суммарная биомасса обоих видов цистозир составляет 88,2%. Весной, летом и осенью на глубине 2 и 5 м доля цистозир по биомассе максимальна (95-99%), зимой она снижается до 76-78%. На глубине 10 м во все сезоны, кроме весны, доля этих видов достоверно ниже (в среднем 80-83% летом и осенью и 63% зимой). Вывод о том, что сезонность сообществ C. crinita в основном проявляется в изменении биомассы C. crinita, был сделан и по результатам работ в Средиземном море (Sales, Ballesteros, 2012). Количество видов в пробе в течение года меняется незначительно (достоверные отличия по количеству видов выявлены только между июнем 2012 г. и маем 2013 г. на глубине 2 м и июнем 2012 г. и августом 2012 г. на глубине 10 м). Таким образом, основные паттерны сезонных изменений сообществ C. crinita северо-восточного побережья Черного моря проявляются на глубинах 2 и 5 м. Сезонность проявляется в основном в изменении биомассы вида-доминанта – C. crinita – которая имеет один сезонный минимум (февраль) и сезонный максимум (май-июнь). Сезонная динамика биомассы C. barbata в целом аналогична динамике C. crinita, однако достоверна только на глубине 2 м. Сезонная динамика сообществ на глубине 10 м для большинства исследованных параметров не выявлена.
Список литературы 1. Афанасьев Д.Ф., Корпакова И.Г. Макрофитобентос российского Азово-Черноморья – Ростов-н/Д: ФГУП АзНИИРХ, 2008. – 292 с. 2. Афанасьев Д.Ф., Абдуллин Ш.Р., Середа М.М. Эколого-флористическая классификация донной растительности российского шельфа Черного моря // Известия Самарского научного центра Российской академии наук – 2012. – Т. 14, №1(4), – C. 963-966. 3. Воронов А. Г. Геоботаника. М., 1973. – 384 с. 4. Громов В. В. Методика подводных фитоценотических исследований // Гидробиологические исследования северо-восточной части Черного моря. Ростов-на-Дону, 1973. – С. 69-72. 5. Громов В.В. Донная растительность верхних отделов шельфа южных морей России: Автореф. дис. … докт. биол. наук. – СПб.: БИН РАН, 1998. – 50 с. 6. Камнев А.Н. Структура и функции бурых водорослей. – М.: МГУ, 1989. – 200 с. 7. Калугина-Гутник А.А. Фитобентос Черного моря. – Киев, 1975. – 246 с. 8. Мильчакова Н.А. О новых видах макрофитов флоры Черного моря // Экология моря. 2002а. № 62. C. 19–24. 9. Мильчакова Н. А. Бурые водоросли Черного моря: систематический состав и распространение // Альгология. 2002b. № 3. С. 324-337. 10. Ballesteros E. 1990a. Structure and dynamics of the Cystoseira caespitosa Sauvageau (Fucales, Phaeophyceae) community in the North-Western Mediterranean // Scientia Marina. 54. P. 155-168. 11. Ballesteros E. 1990b. Structure and dynamics of the community of Cystoseira zosteroides (Turner) C. Agardh (Fucales, Phaeophyceae) in the north-western Mediterranean // Scientia Marina 54. P. 217-229. 12. Ballesteros E. Els vegetals i la zonació litoral: espècies, comunitats i factors que influeixen en la seva distribució. Arxius de la Secció de Ciències, Institut d’Estudis Catalans. – Barcelona, 1992. – 613 pp. 13. Draisma S., Ballesteros E., Rousseau F., Thibaut T. DNA sequence data demonstrate the polyphyly of the genus Cystoseira and other Sargassaceae genera Phaeophyceae // J. Phycol. 2010. 46(6). P. 1329-1345. 14. Giaccone G. Contributo allo studio dei popolamenti algali del basso Tirreno // Ann. Univ. Ferrara, 1971. 4. P. 17-43. 15. Giaccone G. 1973. Écologie et chorologie des Cystoseira de Méditerranée // Rapp. Com. Int. mer Médit. 22. P. 49-50. 16. Giaccone G., Alongi G., Cossu A. V. L., Di Geronimo R., Serio D. 1993. La vegetazione marina bentonica del Mediterraneo: 1.: sopralitorale e mesolitorale: proposte di aggiornamento // Bollettino dell'Accademia Gioenia di scienze naturali. Vol. 26 (341). P. 245-291. 17. Giaccone G., Alongi G., Pizzuto F., Cossu A. La vegetazione marina bentonica fotofila del Mediterraneo: II. Infralitorale e Circalitorale. Proposte di aggiornamento // Boll. Accad. Gioenia Sci. Nat. Catania. 1994. 27. P. 111-157. 18. Giaccone G., Pignatti S. Studi sulla produttività primaria del fitobentos nel Golfo di Trieste. II. La vegetazione del Golfo di Trieste // Nova Thalassia, 1967. 3 (2). P. 1-28. 19. Molinier R. Etudes des biocénoses marines du Cap Cors. Thèse Doctorat. – Université de Marseille. – 1958. 20. Montesanto B., Panayotidis P. The Cystoseira spp. Communities from the Aegean Sea (NE Mediterranean) // Mediterranean Marine Science. 2001, Vol. 2/1. P. 57-67. 21. Panayotidis P., Orfanidis S., Tsiamis Κ. Cystoseira crinita community in the Aegean Sea // Rapp. Comm. int. Mer Medit. 2007. 38. P. 570. 22. Pignatti S. Associazioni di alghe marine sulla costa veneziana // Mem. Ist. Ven. Sci. Lett. Arti. Venezia. 1962. 33. P. 3-131. 23. Pizzuto F. On the structure, typology and periodism of a Cystoseira brachycarpa J. Agardh emend. Giaccone community and of a Cystoseira crinita Duby community from the eastern coast of Sicily (Mediterranean Sea) // Plant Biosyst. 1999. 133. P. 15-35. 24. Ribera M.A., Gómez-Garreta A., Gallardo T., Cormaci M., Furnari G., Giaccone G. Check-list of Mediterranean Seaweeds. I. Fucophyceae (Warming 1884) // Botanica Marina. 1992. 35. P. 109-130. 25. Sales M., Ballesteros E. Shallow Cystoseira (Fucales: Ochrophyta) assemblages thriving in sheltered areas from Menorca (NW Mediterranean): relationships with environmental factors and anthropogenic pressures // Estuar. Coast. Shelf Sci. 2009. 84. P. 476-482. 26. Sales M., Ballesteros E. Seasonal dynamics and annual production of Cystoseira crinita (Fucales: Ochrophyta)-dominated assemblages from the northwestern Mediterranean // Scientia Marina, 76(2), June 2012, p. 391-401. 27. Tsiamis Κ., Orfanidis S., Diapoulis A., Panayotidis P., Siakavara A., Τsiagga Ε., Varkitzi I. Cystoseira crinita community under reference conditions (WFD 2000/60/EE) in the Aegean Sea (Eastern Mediterranean) // 8th Hellenic Symposium of Oceanography, Thessaloniki 2006. p. 319-323. Статья поступила в редакцию 6.12.2015
Communities of brown algae Cystoseira from the north-eastern part of the Black Sea D.F. Afanasyev, A.N. Kamnev, I.V. Stukolova Communities of brown algae Cystoseira from the north-eastern part of the Black Sea have been studied. C. crinita community structure at depths of 2, 5 and 10 m have been revealed. It is found that seasonal fluctuations are manifested mainly in biomass changes of the dominant species C. crinita. Seasonal dynamics of the Cystoseira barbata biomass is similar to the dynamics of C. crinita, however, it reveals itself only at a depth of 2 m. Seasonal dynamics of the communities at a depth of 10 m has not been revealed for the majority of the parameters studied. Key words: Black Sea; phytobenthos; Cystoseira; structure; biomass; total projective cover.
Об авторах Афанасьев Дмитрий Федорович – Afanasyev Dmitry F. кандидат биологических наук Dafanas@mail.ru Камнев Александр Николаевич – Kamnev Alexander N. доктор биологических наук dr.kamnev@mail.ru Стуколова Ирина Владимировна – Stukolova Irina V. ФГБОУ ВПО «Московский государственный университет им.М.В.Ломоносова», Москва, Россия (Lomonosov Moscow State University, Moscow, Russia), Биологический факультет, кафедра физиологии растений iri_sk@inbox.ru Корреспондентский адрес: Россия, 344002, г. Ростов-на-Дону, ул. Береговая, 21 в, АзНИИРХ; тел. (863)-2624197.
ССЫЛКА НА СТАТЬЮ: Афанасьев Д.Ф. , Камнев А.Н. , Стуколова И.В. Сообщества бурых водорослей рода Cystoseira северо-восточной части Черного моря // Вопросы современной альгологии. 2015. № 3 (10). URL: http://algology.ru/847 Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно. При перепечатке ссылка на сайт обязательна
На ГЛАВНУЮ
|
|||
|
| ||