
|
|
|
|
Экологическая физиология водных фототрофных организмов Часть 1. Водные оксигенные фототрофы 1.2. Особенности условий и ресурсов среды обитания водных оксигенных фототрофов
|
|||||||||||||||||
|
Энергетические величины (в скобках – синонимы) |
|
|
Энергия
излучения |
Дж |
|
Поток
излучения |
Вт |
|
Энергетическая
освещенность |
Вт / м2, кал/см2 × мин |
|
Энергетическая экспозиция |
Дж × м-2 |
|
Световая освещенность |
Лк, фот |
|
Квантовое излучение |
мкмоль/м2 × с |
Для монохроматического света излучение и плотность потока фотонов являются связанными характеристиками.
1 мкмоль/м2 × с = (119,7 / l) Вт/м2
где λ – длина волны света в нанометрах.
Для света с более широким спектральным диапазоном переход от излучения к ППФ требует интегрирования значений ППФ, полученной по всей интересующей области спектра. Хотя в целом невозможно точно соотнести световую освещенность, выраженную в люксах, с энергией света или плотностью потока фотонов, неплохим приближением (для большинства источников белого света), можно использовать для этого соотношение:
1 Вт/м2 = 5 мкмоль/м2 = 250 лк.
Проходя через атмосферу, на нашу Землю падают как прямые солнечные лучи – прямая солнечная радиация, так и солнечные лучи, которые во время прохождения через атмосферу и облака становятся рассеянными, т.е. рассеянная радиация. Более того, солнечные излучения достигают Земли в основном в измененном виде, что благоприятно сказывается на жизни Планеты. Так, часть ультрафиолетовых лучей поглощается озоновым слоем, что снимает риск ионизации в молекулах живых систем. Тепловые инфракрасные излучения поглощаются присутствующими в атмосфере парами воды и двуокисью углерода, вследствие чего температура на поверхности нашей Планеты поддерживается на том уровне, который необходим для жизнедеятельности живых систем. Проходя через атмосферу, поверхности Земли достигают в основном солнечные лучи видимой и инфракрасной областей всего солнечного спектра. Именно эти виды солнечных излучений составляют основу энергетики всех живых организмов, именно эту энергию усваивают и в процессе фотосинтеза фототрофные организмы.
В итоге после прохождения через атмосферу и рассеивание на поверхность Земли ежегодно поступает 2·1017 Вт, примерно 100 000 калорий (418 400 джоулей) энергии, приходящихся на каждый квадратный сантиметр. Освещенность Земли, создаваемая Солнцем, когда оно находится в зените, равняется около 100 тыс. лк. Около одной трети этой энергии расходуется на испарение водных масс Земли, а остальная энергия может быть использована в процессе фотосинтеза для построения органического вещества Планеты. Интересно отметить, что эффективность использования солнечной энергии фототрофными организмами в процессе фотосинтеза невысока и, по мнению разных исследователей, колеблется от 0,1 до нескольких процентов. Тем не менее, этого достаточно для поддержания жизни, т.е. термодинамического баланса на нашей Планете.
Как уже говорилось выше, в процессе термодинамических реакций, происходящих на Солнце, высвобождаемая энергия дифференцируется на целый спектр излучений, имеющих различные длины волн, а соответственно обладающих разной энергией. Основная часть фототрофных организмов растительного типа (голофитных организмов), за исключением ряда фотосинтезирующих бактерий, воспринимает и использует в энергетических процессах фотосинтеза видимую часть спектра солнечного излучения – от 400 до 700 нм. Некоторые фотоситезирующие бактерии могут использовать малоэнергетические инфракрасные лучи.
Солнечная энергия излучается в виде определенных порций, т.е. квантов (или фотонов), а их энергия прямо пропорциональна частоте излучений:
E = h × n,
где E – энергия кванта света, h – постоянная Планка, n – частота излучения.
Так как скорость распространения всех видов солнечного излучения равна скорости света 3×1010 см/с, а произведение частоты излучения на длину волны равно скорости света, то частота излучения может быть определена по длине волны и наоборот.
C = l × n,
где С – скорость света, l – длина волны, n – частота излучения.
Таким образом, чем больше частота, тем меньше длина волны, тем выше энергия квантов; т.е. квант синего света обладает большей энергией, чем квант красного света.
При контакте с молекулой квант может быть поглощен ею и, обладая определенной энергией, способен перевести молекулу в «возбужденное состояние», что позволяет молекуле вступить в реакцию, которая ранее для этой молекулы была невозможной.
Кванты рентгеновских и ультрафиолетовых лучей могут ионизировать молекулу, выбивая из атомов электроны. Кванты видимой области спектра обладают меньшей энергией, но при поглощении специальными фотосинтетическими пигментами способны «возбуждать» эти молекулы и запускать фотосинтетический процесс. Кванты инфракрасной области спектра в основном несут тепло и могут вызывать перестройки лишь у ряда молекул, требующих очень незначительного количества энергии.
Две трети поверхности нашей планеты занимает Мировой океан, а также всевозможные континентальные водоемы. Поэтому до 70% от всего количества солнечной энергии, поступающей на Землю, падает на водную поверхность. Вода по сравнению с воздухом менее прозрачная. Более того, чаще всего она насыщена большим количеством взвешенных частиц и живых организмов.
Так как огромная поверхность океана (360×106 км2), в отличие от суши (150×106 км2), практически ничем не затенена, то на нее приходится большое количество прямой солнечной радиации, несущей колоссальные потоки энергии и освещенности. Важно понимать, что в разных широтах в течение года на поверхность воды поступает разное количество солнечной энергии. Так, на 1 см2 в высоких широтах поступает 170-210 кДж, в средних – 330-420, в низких – 580-670 кДж. Та часть потока солнечной энергии, которая может быть использована фототрофами, называется фотосинтетически активной радиацией (ФАР).
Падая на водную поверхность, солнечные лучи отчасти отражаются еще в атмосфере, а частично преломляясь, проникают в водную среду. Кроме того, на степень отражения влияет угол падения солнечных лучей на водную поверхность и степень гидродинамической активности воды. При отвесном попадании лучей на водную поверхность отражение составляет не более 2%. При снижении угла падения лучей до 30º-5º, отражение увеличивается до 25-40% соответственно. При волнении степень отражения также увеличивается. Например, при легком волнении отражение увеличивается до 15%, при сильном – до 30%. По мере прохождения через водную толщу, свет довольно быстро поглощается и рассеивается. При поглощении свет превращается в основном в тепловую и в меньшей степени в химическую энергии, при рассеивании происходит изменение направления потока световых лучей от первоначального в разные стороны (рис. 1).
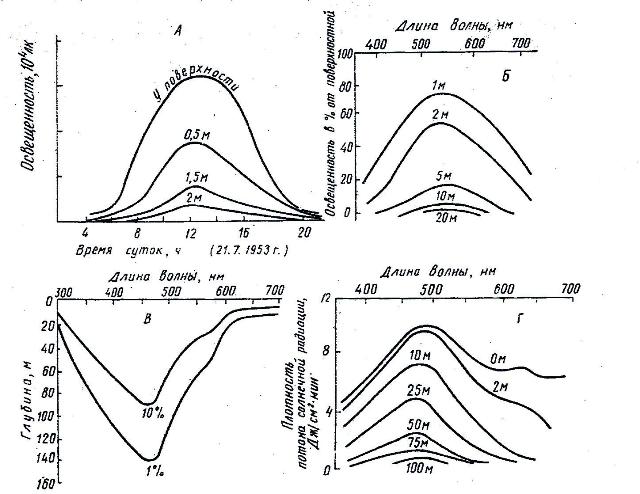
Рис. 1. Проникновение света в толщу воды
А – освещенность разных горизонтов воды Цимлянского водохранилища в течении дня (по Потапов, 1956); Б – освещенность разных горизонтов воды в Нижнем Лунцком озере (прозрачность 8-12 м) в процентах от освещенности поверхностного слоя (по Rutter, 1962); В – глубины, на которых в чистой океанической воде солнечная радиация составляет 1 и 10% от всей падающей на поверхность (по Raymont, 1963); Г – величина солнечной радиации на разных глубинах в Индийском океане (по Raymont, 1963).
Поглощение света описывается законом Бугера-Ламберта-Бэра:
Iz = I0 × e-mz,
где Iz – интенсивность (энергия) световой волны после прохождения ею водного пространства толщиной z, I0 – исходная интенсивность световой волны, m – коэффициент поглощения света, зависящий от длины волны (l).
Так как лучи разной длины волны обладают разной энергией, то их поглощение в водной среде происходит в определенной последовательности. В первую очередь поглощаются инфракрасные (более 820 нм), затем красные (680 нм) и оранжевые (620 нм). Наибольших глубин могут достигать зеленые (520 нм) и синие лучи (460 нм), что влияет на условия фотосинтеза и на поведение гидробионтов (Таблица 1).
Таблица 1. Изменение поглощения
света в чистой воде в зависимости от коэффициента длины волны
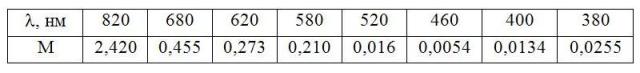
Так как все водоемы содержат большое количество различных взвешенных частиц, то коэффициенты поглощения всех лучей несколько выше.
Как уже говорилось, наряду с поглощением света в толще воды происходит его рассеивание. Ослабление светового потока за счет рассеивания описывается уравнением:
IT = I0 × e-kz,
где k – коэффициент молекулярного рассеивания, зависящий от длины волны лучей (k = 0,000156 × l-4).
Молекулы воды в большей степени рассеивают коротковолновые лучи, а взвешенные в водной среде частицы, напротив, преимущественно длинноволновые лучи, что приводит к рассеиванию света в десятки и сотни раз выше, чем молекулярное.
Суммарное ослабление света за счет его поглощения и рассеивания описывается формулой
IS = I0 × e-(k+m)×z ,
где (k+m) – суммарный коэффициент затухания света.
Так как
все природные водоемы в той или иной степени обогащены всевозможными
взвешенными частицами и гидробионтами, то все они имеют разную прозрачность[7]
и цвет, а это, в свою очередь, также отражается на проникновении
и передаче своей энергии солнечными лучами на различные глубины водоема.
_____________________________
В большинстве рек, прудов и водохранилищ вода имеет минимальную прозрачность и уже на глубину 1 м в большинстве случаев проникает не более 5-10% солнечной энергии, а глубже 2 м от этой энергии остаются десятые доли процента, что составляет 0,015-0,04 Дж/см2×мин. В прозрачных природных озерах солнечные лучи уже могут проникать на глубину до 20-30 м и более, что позволяет поддерживать на этих глубинах энергию солнечной радиации в количестве от 0,04 до 0,004 Дж/см2×мин. Наибольшей прозрачностью отличаются моря, причем самыми прозрачными являются моря южного полушария. В этих морях даже на глубины более 200 м, а в ряде случаев и глубже проникает солнечная радиация, что достаточно для фотосинтетической деятельности гидробионтов.
Несмотря на принципиально другое качество морских вод, прозрачность воды в них, также как и в пресноводных водоемах, может сильно варьировать. Так, например, в Азовском море в период цветения водорослей прозрачность воды может снижаться до 10 см, а в центральных частях океанов, где вода практически не содержит биогенов, необходимых для развития фитопланктона, прозрачность воды может достигать 50 м и более. Максимальная прозрачность – 79 м – установлена в море Уэделла (Антарктида).
С прозрачностью водоема связан и его цвет, который определяется отношением светового потока, выходящего из воды, к потоку, падающему на поверхность. Цвет воды, с одной стороны, зависит от качественных характеристик самого света, в частности от избирательности поглощения и рассеивания солнечных лучей, проникающих в воду под разными углами. С другой стороны, цвет воды влияет на проникновение определенных солнечных лучей вглубь водоема. Чистая вода рассеивает в основном коротковолновые лучи и поэтому она кажется синей, темно-синей, темно-голубой, что говорит о ее органической бедности. В открытом океане прозрачность и цвет определяются в основном количеством планктона и, в частности, фитопланктона, поэтому во время сезонного размножения и интенсивного развития фитопланктона прозрачность воды уменьшается, а цвет становится зеленоватым или зеленым. Аналогичная картина наблюдается с увеличением в воде количества фитопланктона и взвешенных частиц, что характерно по мере приближения к береговой зоне. В этой зоне вода становится не только более зеленой, по сравнению с центральной частью моря или океана, но даже приобретает желтоватые и коричневатые оттенки. Это объясняется тем, что прибрежные воды опресняются стоками рек, богатыми всевозможными взвесями. Кроме того, прибрежные мелководья активно взмучиваются во время штормов.
Таким образом, по цвету воды можно судить о ее чистоте, а также о количестве взвеси и гидробионтов, что, в свою очередь, важно для предварительной оценки энергетического ресурса водоема. Цвет воды водоема нельзя путать с цветом поверхности водоемов. Цвет поверхности зависит от погодных условий и угла зрения на нее.
В соответствии со снижением уровня освещенности по мере увеличения глубины водоемов в них выделяется три зоны. Верхняя толща воды или зона, где освещенность достаточна для энергетических реакций фотосинтеза, называется эвфотической (или просто фотической). По разным источникам фотическая зона различных морей распространяется, по крайней мере, до глубины 200 м, но скорее всего не глубже 300 м. Интересно отметить, что основная масса фитопланктона обитает на глубине 100-150 м. Более того, многие виды фитопланктона обитают исключительно на глубинах 100 м и более. Многие макроводоросли, в том числе даже талломные зеленые, также достаточно широко распространены на больших глубинах. Например, в Атлантике на шельфе штатов Северная и Южная Каролина (США) 25% зеленых водорослей обитает на глубинах более 50 м, на Гавайях обилие зеленых макроводорослей на глубине 90 м выше, чем красных. Более того, на глубине 157 м обнаружена талломная форма зеленой водоросли Johnson-sealinkia profunda. Многие редуцированные, а также корковые красные и зеленые водоросли встречаются и глубже.
Далее простирается дисфотическая, или сумеречная, зона. Глубина этой зоны, в зависимости от моря, может распространяться не менее, чем на 1 км. Считается, что минимальные количества энергии солнечного света, проникающего в дисфотическую зону до глубины 1 км и более (1150 м) могут улавливаться и использоваться, но уже не фототрофами, а глубоководными рыбами и головоногими моллюсками для отдельных стадий их жизненных циклов. Глубинная часть, куда свет не проникает вообще, называется афотической.
Если разделение вышеперечисленных зон провести с учетом их привязки к дну, т.е. к бентали (bentos (греч.) – глубина), то основная масса фототрофных организмов сосредоточена в материковой отмели – шельфе, хотя здесь важно сделать определенные пояснения. До недавнего времени считалось, что шельф ограничен со стороны моря изобатой 200 м, и поэтому неретическая зона (мелководная зона нереста) ограничивалась этой глубиной, что совпадало и с фотической зоной. В настоящее время этот вопрос пересматривается, т.к. Баренцево море представляет собой шельф, но глубины здесь в основном достигают 400 м. Каспийское море в геологическом отношении также является шельфом, а глубины достигают 800 м. В шельфовой зоне Охотского моря преобладают глубины более 1000 м. Поэтому нельзя отождествлять понятия материковая отмель – шельф, неретическая и фотическая зоны. Фотическая зона может быть ограничена меньшей глубиной, а шельф может простираться глубже. Вероятно, правильнее неретическую зону ограничивать глубиной фотической зоны.
В ряде случаев бенталь подразделяют на фиталь и афиталь. В гидробиологической терминологии на фитале размещаются три важные участка-зоны, которые и формируют фотическую зону. Выше уровня приливов расположена часть берега, которая увлажняется брызгами и заплесками во время штормов. Эта зона называется супралитораль (supra (лат.) – выше, сверху; litus, litoreus (лат.) – берег, береговой, прибрежный). Ниже супралиторали лежит зона, которая периодически заливается водой во время приливов. Это литораль. Ниже приливно-отливной зоны лежит сублитораль. Эта зона простирается до нижней бентальной границы обитания фототрофов.
С увеличением глубины количество света снижается и на определенной глубине, называемой компенсационной точкой (или глубиной), энергия света, которую фототрофные организмы используют для фотосинтеза, становится не достаточной для того, чтобы превысить энергетические траты на дыхание (образование кислорода становится равным потреблению). Это, вероятно, с одной стороны, может тормозить распространение фототрофных организмов на большие глубины, а с другой, позволяет использовать фототрофам внешние готовые органические субстраты, т.е. они могут становиться миксотрофами. Проявление способности к гетеротрофному питанию с увеличением глубины отмечалось разными авторами. Считается, что компенсационная точка для большинства морских макрофитов колеблется в диапазоне освещенности от 32 до 320 лк, что соответствует 0,2-0,7% от всей видимой солнечной энергии. Компенсационная точка отражает некую среднюю непрерывную освещенность, необходимую для существования фототрофов (рис. 2).
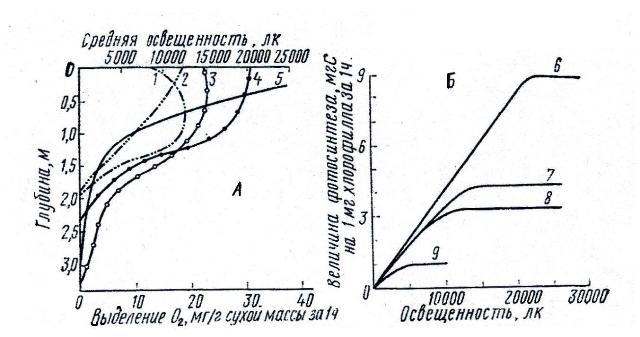
Рис. 2. Зависимость фотосинтеза от освещенности
А – разные растения в одном местообитании (по Потапов,
1956); Б – фитопланктон из разных
местобитаний (по Raymont, 1963):
1 – Utricularia vulgaris, 2 – Ceratophyllum demersum, 3 – Cladophora sp., 4 –
Myriophyllum verticillatum, 5 – освещенность, 6 – тропические поверхностные формы, 7 – тепловодные поверхностные формы, 8 – арктические поверхностные формы, 9 – подповерхностный планктон.
Компенсационная точка зависит от температуры воды. Так, например, компенсационная точка для представителей рода Spirogira при повышении температуры с 5° до 20°С поднимается с 27 до 174 лк, у представителей рода Cladophora – с 63 до 253 лк. Максимальная глубина, на которой освещенность во время наибольшей солнечной радиации соответствует компенсационной точке, превышает по крайней мере в 2,5 раза прозрачность данной области моря. В пресных водоемах нижняя граница компенсационной точки чаще всего не превышает нескольких метров.
Что касается количества света, то высшие водные растения и водоросли часто довольствуются очень слабым освещением, гораздо меньшим, чем наземные растения. Эта теневыносливость определяет ту предельную глубину, на которой проникающий в воду свет обеспечивает им жизнь. Она не превышает, как правило, 15-45 м, хотя представители некоторых видов встречаются на глубинах 150-200 м. Основная масса планктонных организмов в морской воде размещается на глубине 40-70- м, а в некоторых местах они опускаются на глубину 150-200 и более метров. В озерах фитопланктон встречается преимущественно на глубине 10-15 м, а в водах с малой прозрачностью – в самом поверхностном слое толщиной не более 3 м. Считается, что более требовательны к интенсивности освещения зеленые и синезеленые водоросли, а большинство диатомовых, наоборот, избегает ярко освещенного приповерхностного слоя воды.
Учитывая то, что площадь поверхности Океана более чем в два раза больше площади поверхности Земли, при этом суша и вода распределены на поверхности Земного шара неравномерно (в Северном полушарии океаническая поверхность занимает примерно 61%, а суша – 39%; в Южном – примерно 81% и 19% соответственно), можно понимать, что по крайней мере 70% общего количества энергии солнечного излучения, равного 6,3×1020 ккал в год, падает на поверхность морей и океанов, причем основное количество энергии принимают поверхностные воды Южного полушария.
Более того, солнечная энергия проникает на глубины 200-300 м и как толстым энергетическим одеялом покрывает 2/3 поверхности Планеты, обеспечивая таким образом существование фототрофов, производящих живое органическое вещество и O2, поддерживая глобальный геохимический круговорот. Таким образом, зная принципиальную схему солнечных энергетических потоков в Мировом океане, думается, что в ближайшее время это можно будет рационально использовать.
Температура. Температура – один из наиболее важных факторов, обусловливающий нормальную жизнедеятельность высших водных растений и водорослей. В целом для них характерен широкий диапазон температурных условий обитания. Тем не менее, одни виды приурочены исключительно к холодноводным водоемам, другие – к водоемам с теплой водой, а третьи живут в разных условиях, т.е. способны переносить существенные колебания температуры.
Виды,
которые развиваются в сравнительно узких пределах изменения температуры,
называются стенотермными видами. Так, большинство ламинариевых водорослей
и многие виды фукусов – типичные обитатели холодных вод как северного, так и
южного полушария. В то же время близкие родственники фукусов саргассовые
водоросли являются теплолюбивыми и обитают преимущественно в тропиках. В
противоположность им эвритермные[8]
виды встречаются в широком интервале изменения температурного фактора, что
обусловливает их присутствие в водоемах разных географических широт и в разные
сезоны года. Примером
эвритермных водорослей может служить также представитель семейства ламинариевых Macrocystis, ареал
которого простирается с севера и юга до тропиков, вдоль тихоокеанских берегов
Северной и Южной Америки. Среди фитопланктонных организмов также известны и стенотермные, и
эвритермные виды.
_____________________________
Важно отметить, что температурные границы встречаемости видов, определяемые максимальной и минимальной летальными температурами, значительно шире зоны температурного оптимума, в пределах которого жизненные функции осуществляются наиболее активно, что выражается в наибольшей продуктивности данного вида водоросли. Например, диатомовая водоросль Melosira islandica, распространенная в озерном планктоне умеренной и субарктической зон, обычно встречается при температуре воды от 1 до 130С, максимальное же размножение ее происходит в более узком интервале температур – от 6 до 80С.
Поскольку температурные оптимумы у отдельных видов разные, а температура воды изменяется по сезонам, в водоемах происходит периодическая смена одних доминирующих видов водорослей другими, так называемая сезонная сукцессия видов. Иллюстрацией этому служит годовое развитие фитопланктона в озерах умеренно-холодного климата. Зимой в связи с образованием ледяной корки фитопланктон не развивается из-за недостатка освещения. В марте – апреле в довольно большом количестве появляются мелкие жгутиковые Cryptomonas, Chromulina и некоторые другие. В мае бурно развиваются диатомовые водоросли, среди которых преобладают виды Melosira и Diatoma. Холодноводный комплекс диатомей с повышением температуры воды до150С сменяется умеренно-тепловым, в состав которого входят не только диатомовые (Asterionella, Tabellaria и др.), но также хризомонады, зеленые и синезеленые водоросли. При дальнейшем повышении температуры представители зеленых и синезеленых становятся доминирующими. В летнем планктоне диатомеи сравнительно немногочисленны, к их тепловодным видам относятся виды Fragilaria, Melosira granulata. Осенью с понижением температуры до 10-120С снова наблюдается вспышка развития холодноводных видов диатомей.
Двувершинная кривая развития, отмечаемая главным образом для диатомовых водорослей, динофлагеллят и хризомонад, характерна и для морского планктона. В северных морях России при весеннем цветении господствуют обычно виды Chaetoceros, осенью – динофлагелляты.
Следует, однако, отметить, что, несмотря на доминирующее значение температурного режима сезонная сукцессия связана и с рядом других факторов, в частности содержанием азотистых соединений, которых особенно много в воде именно в весенне-осенний период.
Химический состав
водной среды. Важнейшими химическими компонентами
водной среды являются минеральные соли, биогенные элементы[9],
минеральные соединения, органические вещества (растворенные и взвешенные) и
органоминеральные скопления (детрит).
_____________________________
[9]Биогенные элементы – химические элементы, постоянно входящие в состав организмов и необходимые им для жизнедеятельности. К ним относятся кислород, углерод, водород, азот, кальций, калий, фосфор, магний, сера, хлор, натрий.
____________________________
[10] Это деление чаще используют для пресноводных водоемов, прудов и озер.
Естественные воды
довольно резко отличаются между собой по суммарной концентрации солей, или общей
солености. Именно этот фактор обусловливает разделение водоемов и населяющих их
высших растений и водорослей на две большие группы: морские и пресноводные. Морские
воды имеют соленость 35-38‰ (35-38 г соли/л раствора), пресноводные
– с соленостью менее 1‰[11].
Промежуточное положение занимают солоноватые воды (эстуарии больших рек,
лагуны, где морская вода опресняется за счет дождей или поступления пресных вод и т.п.).
_____________________________
Флора высших растений и водорослей пресных вод и морей различна. Типичными обитателями пресных вод являются перечисленные ранее высшие растения: мхи, папоротники, хвощи, покрыто- и голосеменные растения. Среди покрытосеменных, например, достаточно простые нимфейные, лютиковые и высокоорганизованные – колокольчиковые и лилейные. Среди голосеменных болотные кипарисы и сосны. Среди низших фототрофов почти все конъюгаты, большинство вольвоксовых и хлорококковых водорослей. Сифоновые (бриопсидовые), наоборот, являются морскими формами. Подавляющее большинство бурых и красных водорослей также распространено в море. Синезеленые и диатомеи встречаются как в морях, так и в пресных водах. Однако общие виды, которые одинаково хорошо развивались бы и в морской и в пресной водах, редки.
Альгофлора солоноватых вод, очень изменчивых по концентрации и составу солей, представлена эвригалинными формами, которые легко приспосабливаются к большим колебаниям солености. Эти водоросли не отличается большим видовым разнообразием, зато представлены большим числом особей, так как не имеют конкурентов со стороны других организмов.
Для фототрофов наиболее важными из минеральных веществ являются соли азота и фосфора. Соли азота эти растительные организмы могут использовать в любой встречающейся в природе форме. Относительное содержание солей азота в водоемах может меняться, но общее количество данного элемента из года в год остается величиной постоянной, равной примерно 1 мг/л. Содержание фосфора в воде измеряется гораздо меньшими величинами, в среднем от 0,5 мг до тысячных долей мг на 1 л.
Однако во многих естественных водоемах количество азота и фосфора часто невелико, что может тормозить развитие фототрофов. Особенно обеднена в отношении этих элементов открытая, удаленная от берегов часть морских бассейнов. Обогащение таких вод в результате вертикальной циркуляции воды, когда придонные воды поднимаются вверх (явление апвеллинга) в зону фотосинтеза, или за счет приноса питательных веществ речной водой ведет к резкому увеличению продуктивности водорослей.
Железо и кальций, необходимые для питания высших растений и водорослей, используются ими в ионной форме. Содержание этих элементов в разных водоемах сильно варьирует, причем для железа эти колебания выражены в большей степени, что часто зависит от климатических условий.
В водоемах существует естественная связь между содержанием кальция и железа, а также углекислоты и железа. В известковых водах, имеющих щелочную реакцию, соли железа выпадают в осадок. В водах, бедных известью, с нейтральной и кислой реакцией, наоборот, наблюдается повышенное содержание железа в растворе.
Потребности разных видов фоторофов в железе и кальции не одинаковы, а часто даже и прямо противоположны, поэтому накопление этих элементов в воде – решающий фактор в распределении растений. К числу железолюбивых форм, произрастающих в среде с концентрацией Fe 1-3мг/л, относятся многие диатомовые, десмидиевые водоросли, драпарнальдия, хетофора, улотрикс, вошерия и др. В жестких водах с большим количеством кальция (более 25 мг/г) и минимальным количеством железа (менее 0,5 мг/л) обитают кладофора и хара.
Водоросли, обладающие повышенной чувствительностью к железу, такие как Cladophora fracta и Oedogonium capillare, могут использоваться в качестве индикаторов содержания железа в воде. Например, кладофора лучше всего развивается при концентрации Fe 0,2-0,3 мг/л; при повышении же концентрации Fe до 0,5 мг/л ее рост замедляется. Для эдогониума оптимальная концентрация Fe в среде – 0,4-0,5 мг/л, при большей концентрации его развитие останавливается.
Магний, калий и сера также принадлежат к числу необходимых для высших растений и водорослей элементов, но в отличие от вышеуказанных не имеют экологического значения, поскольку практически всегда присутствуют в среде в достаточных количествах.
Кремний содержится в больших количествах в пресных водах, чем в море, причем наиболее бедны этим элементом поверхностные слои морских вод. Поскольку кремний необходим в основном для построения панцирей диатомовых водорослей, его недостаточное содержание в среде обитания отражается на строении панцирей некоторых диатомей: в результате слабого окремнения у морских форм диатомей панцири становятся заметно тоньше. От количества кремния в воде зависит и размножение диатомей: деление клеток диатомовых водорослей идет нормально, если в воде содержится не менее 5 мг Si/л; когда же его содержание снижается до 0,5мг/л, деление прекращается. Отмечено, что весенняя и осенняя вспышки развития диатомей совпадают не только с температурным оптимумом и содержанием основных элементов питания, но и с максимальным количеством кремния в среде.
Водные фототрофы нуждаются не только в биогенных элементах. Не менее важны для их жизни микроэлементы, которые в природных средах присутствуют в достаточных количествах. Например, повышенное содержание марганца в воде стимулирует рост железолюбивых водорослей.
Водные фототрофы испытывают необходимость и в биологически активных соединениях. На диатомовые и некоторые другие водоросли благоприятно действуют, например, витамины В1, В12.
Некоторые высшие водные растения как, например, альдрованда, пузырчатка, а также многие водоросли используют для своего роста и развития экзогенные органические вещества, которые содействуют ускорению их роста и увеличению биомассы. Т.е. водные фототрофы обнаруживают способность к миксотрофному (смешанному) питанию – как за счет фотосинтезируемых, так и за счет поглощаемых из среды органических веществ.
Усвоение фототрофами растворенных органических веществ показано для зеленых, синезеленых, диатомовых и других в опытах с чистыми культурами. При этом обнаружено, что некоторые виды в присутствии органических веществ могут расти и в абсолютной темноте, как типичные сапрофиты. В таких условиях водоросли Pleurococcus, некоторые виды Chlorella не теряют хлорофилла, другие же, например эвгленовые, обесцвечиваются, хотя и продолжают размножаться. При обильном органическом питании клетки теряют хлорофилл и на свету. У ряда бесцветных форм водорослей (Prototheca, Polytoma и др.) такое состояние наследственно закрепилось, и они встречаются в природных водах с большим содержанием органики как сапрофиты.
Из источников углерода в условиях культуры наиболее доступны для водорослей простые сахара и некоторые органические кислоты. Азотистые органические соединения (мочевина, аминокислоты, растворенные белки) одновременно являются источниками и азота, и углерода.
Способность водорослей и высших растений использовать в процессе жизнедеятельности разнообразные группы растворенных веществ обусловливает их участие в очищении водоемов. Автотрофные растений, выделяя кислород, прямо или косвенно содействуют минерализации органических веществ. В результате такой деятельности водорослей и других микроорганизмов загрязненная и часто имеющая тяжелый запах вода превращается в чистую: происходит процесс биологической очистки вод.
Однако «санитарные» способности высших растений и
водорослей не безграничны. Превышение определенного уровня загрязненности
водоемов в результате хозяйственной деятельности человека приводит к нарушению
процессов их естественной самоочистки и соответственно к резко отрицательным
последствиям для их обитателей, в том числе и самих водных фототрофов. Так,
сброс в водоемы сточных вод промышленных предприятий служит причиной токсикации среды, которая влечет за
собой обеднение или полную гибель фитопланктона. Чрезмерное поступление смытых
с сельскохозяйственных площадей веществ или сточных городских вод, сильно
обогащая водоем биогенами[12],
напротив, вызывает бурное развитие фитопланктона. Водоем начинает «цвести»,
причем очень сильно, что сопровождается разложением водорослей и выделением
токсических веществ, которые вызывают гибель не только флоры и фауны самого
водоема, но и животных, использующих такую воду для питья. Умеренное же
поступлении биогенов в природные воды сопровождается повышением продуктивности
сначала фитопланктона, затем зоопланктона, что в конечном итоге благоприятно
отражается на увеличении рыбных запасов.
____________________________
.
Движение воды. Движение воды – течения, прибой, приливы и отливы, вертикальные и горизонтальные циркуляции – можно рассматривать как один из основных факторов, определяющих рост и развитие водных фоторофов. Это связано с тем, что в местах с высокой подвижностью воды за счет ее постоянного обновления водные фоторофные организмы лучше обеспечиваются питательными веществами. Кроме того, в этих зонах происходит происходит более интенсивное удаление автотоксических продуктов метаболизма, что благоприятно сказывается на росте и развитии водорослей. Как и для других экологических факторов, для движения воды существуют верхние и нижние границы, при которых водоросли могут нормально функционировать. В частности, у макроводорослей изменяется форма тела: например, у Fucus distichus из зоны большой волновой активности таллом становится более разветвленным, но при этом его ветви утоньшаются. Пластины ламинариевых становятся более вытянутыми. Кроме того, у ряда водорослей зоны с повышенной активностью воды отмечается утолщении клеточных стенок, что связывают с увеличение механической прочности этих растительных организмов, а также увеличивается прочность их прикрепления к субстрату. Однако слишком мощное движение водных масс может вызвать гибель молодых макроводорослей или истирание талломов взрослых, отрыв их от субстрата и полное разрушение организма.
Субстрат. Существенное значение для развития прикрепленных высших растений и водорослей имеет субстрат, на котором они растут. Экспериментально доказано, что зооспоры обладают хемотаксисом и способны выбирать субстрат. Например, высшие растения предпочитают рыхлые грунты, за исключением Phyllospadix, бурые водоросли твердые субстраты – скалы, камни, металлические сваи; среди красных макроводорослей встречаются как виды, предпочитающие каменистый субстрат, так и виды, развивающиеся интенсивнее на песчаном или илистом дне. А зеленые водоросли заселяют самые разнообразные субстраты, вплоть до растительных организмов.
Обсыхание. Этот фактор влияет на рост и развитие преимущественно морских водорослей, поскольку именно они подвергаются периодическому обсыханию во время отливов. У макрофитов, растущих в приливно-отливной зоне, в отличие от родственных видов, но растущих постоянно погруженными, в значительной степени могут изменяться биохимия, соотношение РНК и ДНК, функциональные характеристики (интенсивность фотосинтеза, темнового дыхания), форма таллома и скорость его роста. Например, скорость роста Fucus vesiculosus верхней литорали, т.е. периодически обсыхающего, ниже, чем у бурых водорослей средней и нижней литорали. А у Fucus spiralis и Porphyra yezoensis фотосинтез и темновое дыхание при небольшой потере воды (сохранение 96-92% воды в тканях) на воздухе даже стимулирует фотосинтез (до 148% по сравнению с полностью оводненными талломами); большее же обезвоживание вдет к снижению фотосинтеза (до 0 при потере 85% воды у Fucus spiralis и 90% воды у Porphyra yezoensi). Микроводоросли литоральной зоны избегают обсыхания, оставаясь под покровом макроводорослей. Микроводоросли пресноводных бассейнов переживают неблагоприятные условия при обсыхании в виде спор.
Ледовый режим. Лед оказывает как положительное, так и отрицательное воздействие на высшие растения и водоросли. Его положительная функция состоит в том, что он «укрывает» их (кроме литоральных видов), создавая константные (по нулевой и положительной температуре) условия, при которых продолжается замедленный рост слоевища с соответствующим циклом размножения. Косвенное положительное влияние льда проявляется и при его весеннем таянии, в результате которого вода обогащается биогенами и происходит подкормка высших растений и водорослей, способствующая повышению темпа роста ряда макро и микрофитов. В течение зимы лед накапливает биогены, (правда, наряду с загрязнителями), впитывая их в себя подобно губке; концентрация этих веществ в нем может в 5-10 раз (иногда даже в сотни раз) превышать их концентрацию в нормальной морской воде.
Отрицательное воздействие льда проявляется в различных формах. Водоросли, растущие на глубине до 0,5 м, испытывают непосредственное влияние льда: они вмерзают в его толщу и во время ледохода «стираются», в связи с чем на этой глубине невозможно развитие крупных многолетних видов макроводорослей. На больших глубинах водоросли подвергаются опосредованному влиянию льда: интенсивность света уменьшается, что может отрицательно сказываться на фотосинтетической активности макрофитов.
Раннее становление льда приводит к повреждению макрофитов: они «срезаются» острыми краями льдин. Механическое повреждение ранним льдом может составлять 50% всей популяции и более. В результате уменьшается численность составляющих фитоценозы видов, понижается общая биомасса, замедляются темпы роста.
Косвенное отрицательное влияние льда на альгофлору связано с аккумуляцией им загрязняющих веществ. В местах скопления сильно загрязненных береговыми наносами медленнотающих ледовых глыб наблюдается более позднее появление сезонных фитоценозов, снижается в 1,5-2 раза темп роста, уменьшается численность, исчезают наиболее чувствительные к загрязнению виды, развиваются другие фитоценозы.
Биотические факторы
Помимо абиотический факторов на рост и развитие высших водных растений и водорослей действуют биотические факторы - разнородные взаимоотношения между особями одной или разных популяций фототрофовных организмов, а также взаимодействия отдельных групп фототрофов с другими растениями и животными. Эти взаимоотношения имеют следующие формы: симбиоз и мутуализм, комменсализм и аменсализм, хищничество и паразитизм, конкуренция и нейтрализм.
При совместном произрастании высших
растений и водорослей наиболее часто встречающейся формой взаимодействия
является конкуренция. В конкурентной борьбе более
быстрорастущие организмы чисто механически вытесняют медленнорастущие. А в синузиях[13]
представители одного вида воздействуют на
представителей соседствующего вида с помощью выделения определенных химических веществ, тормозящих или полностью
подавляющих рост последних. Например, при проращивании зооспор ламинариевых
водорослей в искусственных условиях проростки из них не развиваются, если в
сосудах с зооспорами находятся части талломов аскофиллума.
____________________________
Однако известны факты и положительного
взаимодействия водорослей, в частности с высшими растениями, а именно
стимулирующее действие прижизненных выделений водорослей на корни высших растений
в почвенных экосистемах – консорциях[14] .
____________________________
[14] Консорция – минимальная структурная единица биоценоза, состоящая из разнородных организмов, которые в течение всей жизни или отдельных периодов находятся в тесных и полезных контактных отношениях друг с другом.
Поскольку водоросли служат пищей различным животным, разрастание их нередко прямо зависит от численности последних. Так, было установлено, что площади, занятые промысловыми бентосными водорослями у берегов Канады, сокращаются из-за выедания их морскими ежами. Регулировать численность этих животных, а следовательно, препятствовать истреблению зарослей макрофитов возможно косвенным путем: сокращением вылова в этих местах омаров, питающихся морскими ежами.

КОНТАКТЫ
Email: info@algology.ru
Изготовление интернет сайта
5Dmedia
ЛИЦЕНЗИЯ
Эл N ФС 77-22222 от 01 ноября 2005г.
ISSN 2311-0147