
|
|
|
|
29 ноября - 29 декабря 2015 г. Исследование влияния различных токсикантов на популяции микроводорослей и инактивация этих соединений
Даллакян Г.А. Genaris A. Dallakyan
Московский Государственный Университет им. М. В. Ломоносова,
УДК 574.583:582.263:574.63
Показано, что различные токсиканты подавляют фотосинтетический аппарат, скорость прироста численности водорослей и удельную скорость выделения кислорода. Эффективность защиты от токсиканта зависит от способов обработки питательной среды шунгитом – предварительной обработки в течение суток среды шунгитом или постоянного присутствия шунгита в среде. При этом шунгит инактивирует токсическое действие как синглетного кислорода, так и тяжелых металлов. Ключевые слова: водоросли; бихромат калия; сульфат кадия; шунгит; флуоресценция; эффективность фотосинтеза.
Введение Микроводоросли являются главными продуцентами в водоемах и первичными мишенями для различных вредных веществ, поступающих в водные экосистемы. Химический состав шунгита исследуется достаточно интенсивно (Каленин, 2008), однако влияние на биологические объекты исследовано мало, хотя этот вопрос очень важный, поскольку в последние годы идет очень интенсивная продажа шунгита для лечения различных недугов человека. Показано, что в проходящей через шунгит воде быстро окисляются органические молекулы и свободные радикалы. При этом содержание в ней бактерий снижается в десятки раз (Voeikov, 2006). Важную роль в очистке воды от загрязняющих веществ и стабилизации энергетического баланса воды играют абсорбционные свойства шунгита и фуллереноподобные вещества. Ряд авторов предполагают, что фуллерены, входящие в состав шунгита, изменяют окислительно-восстановительный потенциал среды и структуру воды, тем самым способствуют очистке воды от вредных для здоровья человека веществ (Пиотровский и др., 2007). Поэтому в нашей работе исследованы различные способы очистки питательной среды для водорослей, поскольку в способах очистки воды остается много неясного. Возможно, питательную среду для водорослей можно «активировать» шунгитом в течение определенного времени (например, в течение суток), а затем использовать ее для защиты от токсиканта без присутствия шунгита. Материал и методы Объектом исследования была альгологически чистая культура Scenedesmus quadricauda (Turp.) Bréb. Водоросли выращивали в конических колбах объемом 100 мл в среде Успенского №1 при температуре 25°С и круглосуточном освещении 15 мкмоль квантов м-2с-1. В экспериментах использовали шунгит с Зажогинского месторождения от компании «Арго». Генератором синглетного кислорода служил фотосенсибилизатор эозин (динатриевая соль эозина C20H8Br4Na2O5) в количестве 5 мгл. Токсиканты бихромат калия в концентрации 3 мг/л, сульфат кадмия в концентрации 1,5 мг/л, эозин и шунгит 100г/л добавляли в среду однократно на 3-й день после посева культуры. Шунгит предварительно обрабатывали согласно инструкции изготовителя с учетом специфики выращивания водорослей. Численность клеток подсчитывали в камере Горяева под световым микроскопом. Определение живых и мертвых клеток в культуре осуществляли с помощью люминесцентного микроскопа Carl Zeiss Axioscop 2 FS Plus в проходящем свете. При облучении объекта короткими сине-фиолетовыми лучами получали длинноволновое видимое свечение объекта. Живые клетки имели ярко-красное свечение, а мертвые – зеленое. Интенсивность флуоресценции хлорофилла и кинетика индукции флуоресценции рассчитывали по показателям Fo и Fm, которые измеряли на приборе «МЕГА-25» (Погосян и др., 2009). Результаты и обсуждение Показано, что скорость прироста популяции водорослей больше в присутствии шунгита в среде. При этом в питательной среде присутствовал эозин. Присутствие в среде только эозина оказывает наименьшую скорость прироста популяции. Ранее в наших опытах было показано, что шунгит инактивирует токсическое действие фотосенсибилизатора (Даллакян, 2014). В этих работах выдвигались гипотезы, объясняющие действие шунгита. Возможно ли «активировать» среду шунгитом после обработки ее в течение фиксированного времени, а затем вырастить водоросли в обработанной среде без шунгита, чтобы инактивирующие свойства среды сохранились? В пробах, обработанных шунгитом, и в контроле скорость прироста популяции находится в пределах ошибки эксперимента. Таким образом, в среде, предварительно обработанной шунгитом, до 10-ых суток прирост культуры сравним с контролем, далее прирост резко снижается. По-видимому, соединения, растворенные в среде во время обработки шунгитом, проводят стабилизацию питательной среды, однако через какое-то время их концентрация в значительной степени уменьшается. В активированных средах защитные свойства шунгита от токсикантов сохраняются частично. Возможно, эти соединения в первые три дня инактивируют действие эозина. Далее их количества не хватает для стабилизации редокс-потенциала среды. На рис.1 представлены результаты исследования жизнеспособности клеток S. quadricauda, оцененной с помощью метода люминесцентной микроскопии. В присутствии 3 мг/л бихромата калия количество живых клеток в культуре со временем постепенно уменьшалось, а доля мертвых – соответственно увеличивалась (рис. 1, а). В присутствии шунгита доля живых клеток в культуре была на уровне контроля.
Рис.1. Доля живых клеток в культуре S. quadricauda (в % от общего количества) в присутствии шунгита 100 г/л с бихроматом калия, 3 мг/л (а) и сульфатом кадмия, 1,5 мг/л (б).
При комбинированном действии бихромата калия и шунгита доля живых клеток составляла 98-99% на протяжении всего эксперимента, и к концу эксперимента была выше уровня контроля. Полученные результаты свидетельствуют о снижении токсического действия бихромата калия на культуру S. quadricauda в присутствии шунгита. В присутствии 1,5 мг/л сульфата кадмия количество живых клеток культуры со временем постепенно уменьшалось и к 15 суткам их доля составляла около 4%, а доля мертвых – соответственно со временем увеличивалась (рис. 2, б). В присутствии шунгита доля живых клеток в культуре была на уровне или выше уровне контроля. При комбинированном действии сульфата кадмия и шунгита доля живых клеток составляла 98-99% на протяжении всего эксперимента и была близка к контролю. Полученные результаты также свидетельствуют о снижении токсического действия сульфата кадмия на культуру S. quadricauda в присутствии шунгита. На рис. 2 представлены данные изменения фотохимического квантового выхода ФСII или эффективность фотосинтеза φ (%). Во всех случаях в присутствии шунгита или при одновременном присутствии в среде шунгита и токсиканта (как бихромата калия, так и сульфата кадмия) эффективность фотосинтеза была близкой к уровню контроля. Наименьшие значения величины эффективности фотосинтеза были в пробах, где присутствовал в среде только бихромат калия или сульфат кадмия, причем в последнем случае эти значения были самыми низкими. Данные по эффективности фотосинтеза хорошо согласуются с данными о доле живых клеток в культуре (рис. 1). Близкие значения (в пределах ошибки измерения) эффективности фотосинтеза после 11-х суток роста водорослей во всех пробах можно объяснить адаптацией культуры в среде. Клетки с низкой эффективностью фотосинтеза элиминировались и в популяции оставались только клетки с высокой эффективностью фотосинтеза.
Рис.2. Эффективность фотосинтеза (в %) S. quadricauda в присутствии шунгита 100 г/л с бихроматом калия 3 мг/л (а) и сульфатом кадмия 1,5 мг/л (б).
Ранее (Даллакян, 2014) было показано, что шунгит защищает популяцию клеток S. quadricauda от повреждающего действия фотодинамических красителей. При этом шунгит дезактивирует синглетный кислород, образующийся от красителя, и тем самым защищает водоросли от его токсического действия. Возможно, в этом процессе фуллереноподобные соединения, входящие в состав шунгита, проявляют себя как антиоксиданты (Andrievsky et al, 2009). В присутствии же бихромата калия (Cr+6) в среде синглетный кислород не образуется. Поскольку соединения шестивалентного хрома более токсичны, чем трехвалентного, одним из возможных механизмов защиты популяции водоросли S. quadricauda от токсического действия бихромата калия в присутствии шунгита является переход шестивалентного хрома в менее токсичный трехвалентный: K2Cr2+6O7 + Sh →Cr2+3Sh, где Sh – активные соединения, входящие в состав шунгита (ионы переменной валентности, фуллерены и др. соединения), восстанавливающие бихромат калия и снижающие его токсичность. Не исключается также возможность образования комплекса K2Cr2+6O7 с шунгитом, который становится менее токсичным для микроводоросли. Шунгит восстанавливает различные окислители, сенсибилизаторы и другие соединения, входящие в состав метаболитов, переводя их в неактивное состояние. Таким свойством обладают фуллерены, входящие в состав шунгита. Фуллерен в составе шунгита находится в виде особых, полярных, донорно-акцепторных комплексов с другими химическими соединениями. При этом в шунгитах в основном присутствует фуллерен С60, и составляет около 0,04 мас.% (Buseck et аl., 1992). Значительное улучшение роста S. guadricauda при комбинированном действии бихромата калия и шунгита, а также сульфата кадмия и шунгита по сравнению с ее ростом в присутствии токсиканта без шунгита, возможно связано с тем, что шунгитовые соединения образуют комплекс с бихроматом калия или сульфатом кадмия и инактивируют их действие. Кроме того, фуллереноподобные соединения изменяют свойства воды. Шунгитовые фуллерены плохо растворяются в воде, однако при настаивании в воде в течении нескольких часов вокруг каждого фуллерена образуется многослойная оболочка из молекул воды, которую называют структурированной водой (Пиотровский и др., 2007), что создает условия для благоприятного роста культуры. Возможно также, что во время роста культуры, когда количество экзометаболитов в среде увеличивается, шунгит выступает как сорбент метаболитов, снижая их ингибирующее действие. Таким образом, бихромат калия и сульфат кадмия оказывают выраженное токсическое действие на культуру микроводоросли Scenedesmus quadricauda по физиологическим показателям и доле живых и мертвых клеток. При комбинированном действии шунгита и бихромата калия или сульфата кадмия шунгит инактивирует их токсическое действие, при этом механизм действия шунгита неспецифичен, что указывает на его возможное использование как универсального средства для очистки воды от различных загрязняющих веществ. . Список литературы 1. Andrievsky G.V., Bruskov V.I., Tykhomyrov A.A., Gudkov S.V. Peculiarities of the antioxidant and radioprotective effects of hydrated C60 fullerene nanostuctures in vitro and in vivo // Free Radical Biolo Med. 2009. V. 47. P. 786–793. 2. Buseck Peter R., Tsipursky Semeon J.; Hettich Robert. Fullerenes from the geological environment // Science. 1992: V. 257. № 5067. P. 215-217. 3. Voeikov V.L. Reactive oxygen species (ROS); pathogens or sources or sources of vital energy? Part 1.ROS in normal pathologic physiology of living systems // Journal of Alternative and Complementari Medicine Mar. 2006. V.12. № 2. P. 111-118. 4. Даллакян Г.А. Рост популяции микроводорослей в зависимости от концентрации эозина в питательной среде // Вода: химия и экология. 2014. № 6. С.117-121. 5. Каленин Ю.К. Экологический потенциал шунгита // Наука в России. Москва, 2008. № 6. С. 39-44. 6. Пиотровский Л.Б., Еропкин М.Ю., Еропкина Е.М., Думпис М.А., Киселев О.И. Механизмы биологического действия фуллеренов – зависимость от агрегатного состояния. // Психофармакологии. Биол. наркол. 2007. Т. 7. №. 2. С. 1548-1554 7. Погосян С.И., Казимирко Ю.В., Гальчук С.В., Конюхов И.В., Рубин А.Б. Применение флуориметра «МЕГА-25» для определения количества фитопланктона и оценки состояния его фотосинтетического аппарата // Вода; химия и экология. 2009. № 6. С. 34-40. Статья поступила в редакцию 10.12.2015
Study the impact of various pollutants on populations of algae and inactivation of these compounds Genaris A. Dallakyan It is shown that various toxicants inhibit photosynthetic apparatus, the rate of population growth of algae, and a specific rate of oxygen. The effectiveness of anti-toxicant depends on the method of processing the culture medium with shungite. These methods are 24 hour preliminary the culture processing with shungite or shungite constant presence in the culture. Thus shungite inactivates the toxic effect as a singlet oxygen, and heavy metals. Keywords: algae; potassium dichromate; potassium sulfate; shungite; fluorescence; photosynthetic efficiency.
Об авторе Даллакян Генарис Арменакович - Dallakyan G.A. кандидат биологических наук honaris@bk.ru Корреспондентский адрес: Россия, 119991, Москва, Ленинские горы, Московский Государственный Университет им.М.В.Ломоносова, д. 1, стр. 12, Биологический ф-т, каф. гидробиологии; тел.: (495) 939-27-73.
ССЫЛКА НА СТАТЬЮ: Даллакян Г.А. Исследование влияния различных токсикантов на популяции микроводорослей и инактивация этих соединений // Вопросы современной альгологии. 2016. № 1 (11). URL: http://algology.ru/964 Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно.
При перепечатке ссылка на сайт обязательна
На ГЛАВНУЮ
|
|||
|
| ||
 а)
а)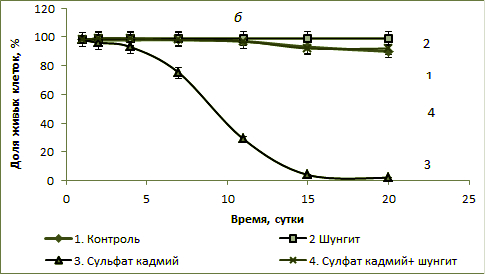 б)
б)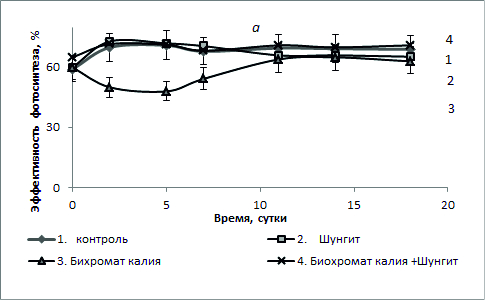 а)
а)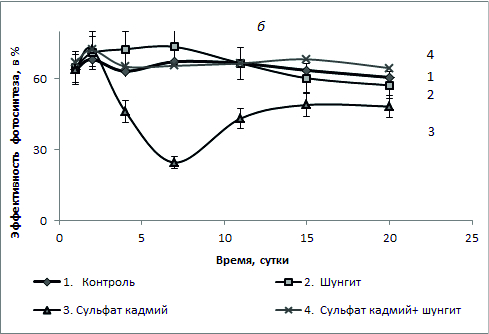 б)
б)