|
|
|
|
29 ноября - 29 декабря 2015 г. Ультраструктурное исследование водорослевого слоя листоватых лишайников
Власова Т.А. Tatyana A. Vlasova
Московский Государственный Университет им. М. В. Ломоносова,
УДК 57.012.4:581.557::582[29+263]
Ультраструктуру водорослевых слоев листоватых лишайников родов Peltigera, Parmelia, Cetraria, Xanthoria исследовали методом сканирующей электронной микроскопии. Было обнаружено, что характер распределения водорослей-фикобионтов в талломах лишайников заметно варьировал. Результаты обсуждаются. Ключевые слова: лишайники; микобионты; фикобионты; Peltigera aphthosa; Parmelia physodes; Cetraria islandica; Xanthoria parietina; электронная микроскопия; ультраструктура.
Введение При изучении лишайникового симбиоза большой интерес представляет выяснение значения каждого симбионта в их сосуществовании и взаимного влияния фикобионта и микобионта. Хотя основную массу таллома и соответственно его форму определяет гриб, но значение водорослевого компонента также очень велико. Именно водоросль-фикобионт, осуществляя фотосинтез, в большой степени снабжает гриб-микобионт органическими веществами, благодаря чему многие лишайники способны расти на самых бедных субстратах. Известно, что фикобионт и микобионт способны к полной делихенизации, то есть могут быть выделены в чистые культуры и расти по отдельности, причем это состояние обратимо: оба симбионта, если их объединить, могут вновь перейти в лихенизированное состояние (Fox,1967; Brunauer, Stocker-Wörgötter, 2005). Под действием неблагоприятных факторов, нарушающих взаимосвязь симбионтов, может возникнуть делихенизация, приводящая к гибели части таллома или даже всего организма. Однако возможна обратимость и подобного процесса в случае изменения условий на более благоприятные условия. В таком случае фикобионт возвращается в нормальное состояние первым, стимулируя репарацию и микобионта (Плакунова, Власова, 1985). Исследование газообмена и ультраструктуры фикобионта при выращивании его в различных условиях позволяют заключить, что изменчивость ультраструктуры фикобионта отражает и его метаболическую подвижность, что способствует выживанию всего организма при изменении условий внешней среды (Власова, Гордонова, Плакунова, 1991). Все это указывает на большое значение фикобионта в общем метаболизме такого симбиотического организма, как лишайник. Целью настоящей работы являлось изучение ультраструктуры водорослевых слоев листоватых, то есть гетеромерных лишайников, фикобионтами которых служат одноклеточные зеленые водоросли (за исключением Peltigera aphthosa, трехкомпонентного лишайника, содержащего в талломе зеленый фикобионт Coccomyxa, а в цефалодиях – цианобионт Nostoc). Талломы лишайников Peltigera aphthosa, Parmelia physodes, Cetraria islandica, Xanthoria parietina собирали в лесу в окрестностях Беломорской биостанции МГУ на Кольском полуострове. Талломы хранили в замороженном состоянии. Для электронно-микроскопического изучения лишайника брали кусочки талломов размером приблизительно 3-5 мм. Материал фиксировали 2,5%-ным раствором глутарового альдегида в фосфатном буфере pH 7,4 и обезвоживали проведением через ряд растворов этанола возрастающей концентрации и 100%-ный ацетон. Затем кусочки талломов высушивали методом перехода критической точки, монтировали на столиках и напыляли тяжелыми металлами. Подготовленный материал просматривали в электронном микроскопе CamScan.
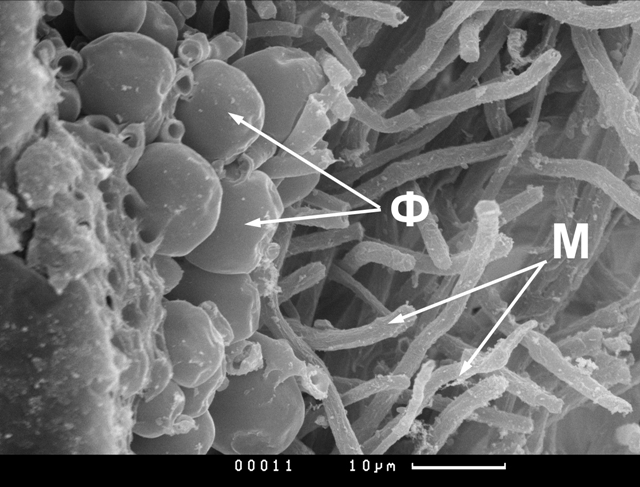
Результаты и обсуждение Изучение срезов лишайниковых талломов, на которых обнаруживались и гифы гриба, и клетки водоролей, выявило значительную вариабельность распределения клеток водорослей в талломах лишайников разных видов. Различия в распределении фикобионта показаны на приведенных ниже электронных микрофотографиях. На рис.1 представлен поперечный срез таллома Peltigera aphthosa. У этого трехкомпонентного лишайника брались участки таллома без цефалодиев, то есть не включающие цианобионт. Видно, что в данном талломе клетки фикобионта Coccomyxa расположены тонким, но компактным, почти сплошным слоем. Заметны довольно близкие контакты клеток фикобионта с гифами микобионта. Клетки водоросли имеют округлую форму, изредка не совсем правильную, в некоторых случаях заметны легкие вмятины, не причиненные контактом с мицелием. Местами встречаются группы тесно сближенных клеток, по всей вероятности, образованные путем деления водорослей. Это соответствует нашим данным, полученным методом просвечивающей электронной микроскопии (Власова, Гордонова, Плакунова, 1991; Власова, 1999). На срезах часто встречаются тетрады водорослевых клеток, образовавшиеся путем деления. Вероятно, интенсивность деления обусловлена определенными условиями.
Рис.1. Ультраструктура водорослевой зоны лишайника Peltigera aphthosa
Иначе выглядит на поперечном срезе строение таллома Parmelia physodes, представленное на рис. 2. В талломе этого лишайника клетки фикобионта Trebouxia, более крупные, чем Coccomyxa, и более правильной формы, занимают значительно большую ширину таллома. Однако они распределены довольно редко, неравномерно, группы разделившихся клеток встречаются заметно реже. Контакты водорослевых клеток с гифами микобионта наблюдаются очень часто, у большинства клеток фикобионта. Сходное с этим лишайником внутреннее строение таллома имеет лишайник Xanthoria parietina, изображение которого здесь не приводится. Интересно отметить, что этот лишайник имеет в качестве фикобионта водоросль рода Trebouxia, как и Parmelia physodes.
Рис.2. Ультраструктура водорослевой зоны лишайника Parmelia physodes.
У лишайника Cetraria islandica (рис.3) фикобионтом является также водоросль рода Trebouxia. Диаметр ее крупных клеток заметно превышает толщину гиф микобионта. Этот фикобионт располагается в талломе в основном довольно компактно, тонким слоем, и активного деления клеток не наблюдается. Имеются случаи довольно тесного контакта мицелия с клетками водоросли. Встречаются небольшие деформации клеток фикобионта, не вызванные контактом с микобионтом.
Рис.3. Ультраструктура водорослевой зоны лишайника Cetraria islandica.
Таким образом, как распределениие фикобионтов в различных лишайниках, так и визуально наблюдаемая интенсивность их взаимодействия с микобионтами различаются довольно значительно. Не исключено, что это определяется свойствами самих водорослей. В пользу этого говорит определенное сходство распределения фикобионтов одного и того же рода Trebouxia в лишайниках Parmelia physodes и Xanthoria parietina. Однако в талломе Cetraria islandica распределение того же фикобионта носит иной характер. Возможно, что основное значение имеет характер симбиотического взаимодействия в данных условиях. Так, показано значение биологически активных вторичных метаболитов, определенных белков и гормонов во взаимодействиях симбионтов лишайников (Воробьев, 2009, Ott, 1993, Stocker- Wörgötter, 2001). Для уточнения этого вопроса требуются дальнейшие исследования. В частности, желательно дополнительно изучить строение талломов лишайников, содержащих те же фикобионты, что и рассмотренные в данной работе.
Список литературы 1. Вайнштейн Е.А., Тахтаджян Е.А. Физиологические изменения у лишайниковой водоросли Trebouxia при культивировании // Физиол. раст. 1981. Т.28, №5. С.1037-1944. 2. Власова Т.А., Гордонова И.К., Плакунова В.Г. Варьирование ультраструктуры Coccomyxa – фотобионта лишайника Peltigera aphthosa в зависимости от условий существования. //Актуальные проблемы экспериментальной лихенологии в СССР. Под ред. Н.С. Голубковой. – Л., 1991. Вып. 1. – С. 66-73. 3. Власова Т.А. Сравнительное исследование ультраструктуры симбионтов лишайника Peltigera aphthosa в разновозрастных участках таллома. // Физиология растений – наука III тысячелетия. Тезисы IV съезда ОФР России. – 1999. – T.1. – С. 109. 4. Власова Т.А., Жевлачкова И.К. Влияние абиотического стресса на ультраструктуру зеленой водоросли Trebouxia 11, фикобионта лишайника Hypogymnia physodes// Физико-химические основы структурно-функциональной организации растений. Годичное собрание ОФР России, Екатеринбург 6-10 октября 2008 г. – Тезисы докладов Международной научной конференции. – С. 111-112. 5. Воробьев Д. В. Ионообменные группы и белки клеточных стенок талломов лишайника Peltigera aphthosa (L.) Wild : Автореферат дис. канд. биол. наук – Москва, 2009 – 21 c. 6. Плакунова В.Г., Власова Т.А. Ультраструктурные изменения при обратимой делихенизации компонентов лишайника Peltigera aphthosa // Изв. АН СССР. Сер. биол. 1985. Т. 3. С. 345-351. 7. Brunauer G., Stocker-Wörgötter E., Culture of lichen fungi for future production of biologically active compounds. // Symbiosis. 2005. 38, №2. P.187-201. 8. Fox C.H. Studies of the cultural physiology of the lichen alga Trebouxia // Physiol. Plantarum. 1967. 20, №1. P. 251-262. 9. Ott S. Analysis of ethylene and ACC in lichens. Protocols in lichenology // Springer Lab. Manuals. 1993. P. 182-195. 10. Stocker-Wörgötter E. Experimental studies of lichen symbiosis : DNA- analyses differentiation and secondary chemistry of selected mycobionts, artificial resynthesis of two-and thripartit symbiosis // Symbiosis. 2001. 30. P. 207-227. Статья поступила в редакцию 20.12.2015 Ultrastructural investigation of algal layers in foliose lichens Tatyana A. Vlasova The ultrastructure of algal layers of foliose lichens (genera Peltigera, Parmelia, Cetraria, Xanthoria) was investigated by mean of scanning electron microscopy. It was found that patterns of phycobiont distribution in the lichen thalli varied significantly. The results are discussed. Key words: lichens; mycobionts; phycobionts; Peltigera aphthosa; Parmelia physodes; Cetraria islandica; Xanthoria parietina; electron microscopy; ultrastructure.
Об авторе Власова Татьяна Анатольевна - Vlasova Tatyana A. кандидат биологических наук tat_vla@list.ru Корреспондентский адрес: Россия, 119991, Москва, Ленинские горы, Московский Государственный Университет им.М.В.Ломоносова, д. 1, стр. 12, Биологический ф-т, каф. физиологии растений; тел.: (495) 939-21-18.
ССЫЛКА НА СТАТЬЮ: Власова Т.А. Ультраструктурное исследование водорослевого слоя листоватых лишайников // Вопросы современной альгологии. 2016. № 1 (11). URL: http://algology.ru/972
Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно. При перепечатке ссылка на сайт обязательна
На ГЛАВНУЮ

|
|||
|
| ||