№ 2 (6) 2014
по Материалам Международной конференции «Экологическая физиология водных фототрофов: распространение, запасы, химический состав и использование»
III Сабининские чтения
29 ноября - 29 декабря 2014 г.
Особенности аминокислотного и жирнокислотного составов
Saccharina japonica Охотского и Японского морей
Features of amino acid and fatty acid compositions
of Saccharina japonica Okhotsk and Japan seas
Гурулёва О.Н., Конева Е.Л., Вишневская Т.И., Кадникова И.А., Павель К.Г., Аминина Н.М.
Olga N. Guruleva, Elena L. Koneva, Tatiana I. Vishnevskaya, Irina A. Kadnikova,
Konstantine G. Pavel', Natalia M. Aminina
Тихоокеанский научно-исследовательский рыбохозяйственный центр (ТИНРО-Центр)
УДК 577.11:582.26(265.53+265.54)
Основными ЖК общих липидов сахарины японской из Японского и Охотского морей являются С16:0, С18:1n-9 жирные кислоты. В водорослях Японского моря также преобладают С18:2n-6, С20:4n-6 ЖК, в сахарине японской Охотского моря – С14:0 ЖК. В водорослях Японского моря определено до 8,9% эйкозапентаеновой кислоты (С20:5n-3). В зависимости от места сбора водорослей доля ПНЖК n-3 серии варьирует от 1,4 до 21,0%, n-6 серии – от 2,0 до 26,9%. Соотношение n-6/ n-3 ПНЖК в водорослях составило 1,0:1 – 2,4:1. На основании проведенных исследований отмечены районы произрастания сахарины японской с высоким содержанием в ней НЖК, МНЖК, ПНЖК, в том числе n-3 серии.
В составе белка исследованных водорослей определено 17 аминокислот, в том числе 7 незаменимых: валин, изолейцин, лейцин, лизин, метионин, треонин, фенилаланин. Отмечено, что в водорослях Охотского моря содержание незаменимых аминокислот выше, чем в сахарине японской Японского моря. Среди заменимых аминокислот преобладают аспарагиновая и глутаминовая кислоты, а также пролин.
Ключевые слова: бурые водоросли, сахарина японская, жирные кислоты, аминокислоты.
Исторически сложилось, что водоросли рассматриваются как источник полисахаридов, маннита и микроэлементов. В тоже время не менее интересны водоросли как продуценты жирных кислоты и аминокислот, обладающих различной биологической активностью.
Жирные кислоты являются одним из главных компонентов в питании человека и животных. Они необходимы для развития и нормального функционирования мозга, зрительной системы, построения клеточных мембран. Наличие в пищевом рационе функционального ингредиента n-3 ПНКЖ обеспечивает снижение сердечно-сосудистых заболеваний (Carroll, Roth, 2002), а также заболеваний связанных с обменом веществ.
Бурые водоросли также синтезируют ряд аминокислот, которые легко усваиваются организмом, участвуют в обменных процессах ЦНС. Белки водорослей обладают повышенной гормоноподобной активностью, полезнее для организма человека, чем белки наземных растений. Это объясняется тем, что в их состав входит моно- и дийодтирозин (Hou et al., 2000).
В литературе представлен аминокислотный и жирнокислотный составы различных видов водорослей (Sanina et al., 2004; Герасименко и др, 2010; Боголицин и др., 2012), показаны различия в накоплении низкомолекулярных метаболитов в водорослях в зависимости от сезона (Honya et al., 1994; Kim et al., 1996; Воскобойников и др., 2005; Sanina et al., 2008; Герасименко и др., 2012). При этом данные по аминокислотному и жирнокислотному составу сахарины японской, произрастающей в Японском море, немногочисленны, а Охотское море не охвачено вовсе.
В Дальневосточном регионе основным объектом промысла является бурая водоросль – Saccharina japonica (сахарина японская). Традиционными районами ее добычи в Японском море считаются прибрежные воды Приморского края (к югу от м. Золотого), в Охотском море – кутовая часть зал. Анива. Общий промысловый запас сахарины японской у побережья Приморского края оценивается в 30 тыс. т (Состояние промысловых…, 2014), в зал. Анива – в 47,8 тыс. т (Евсеева, Репникова, 2010).
Поэтому особый интерес представляет изучение липидного и аминокислотного состава сахарины японской, произрастающей в Дальневосточном регионе. Это позволит выявить различия биохимического состава водорослей и определить наиболее перспективные районы добычи сырья как источника аминокислот и жирных кислот.
Материалы и методы
В качестве объекта исследований использовали слоевища бурой водоросли сахарины японской (Saccharina japonica = Laminaria japonica), которые были добыты в Японском и Охотском морях в сентябре 2013 г. Сбор материала осуществляли водолазным способом в сублиторальной зоне, заготовку проводили в условиях экспедиции путем естественной сушки водорослей на открытом воздухе. Схемы станций сбора водорослей представлены на рисунках 1 и 2.
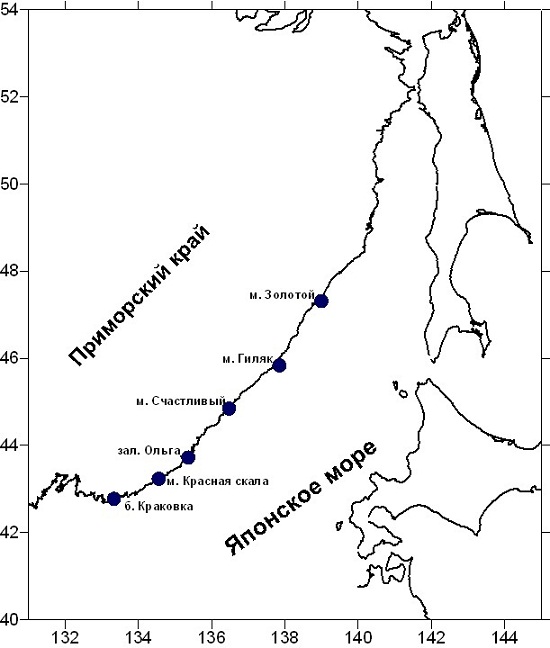
Рис. 1. Схема расположения станций сбора водорослей в Японском море.
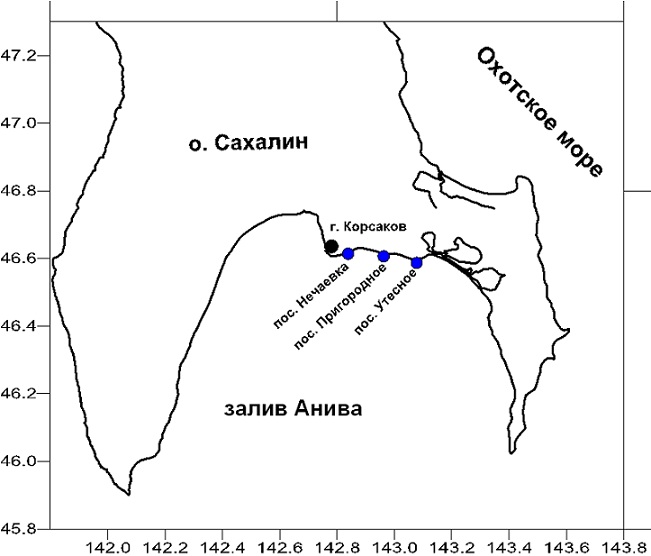
Рис. 2. Схема расположения станций сбора водорослей в зал. Анива (Охотское море).
Для проведения анализов слоевища водорослей измельчали до кусочков размером 0,1-0,5 мм, подготавливали среднюю пробу, которую хранили в банке с притертой пробкой.
Общее содержание азотистых веществ в водорослях определяли микрометодом по Кьельдалю на анализаторе азота «Kjeltec auto» 10 SO Analyzer (Tecator, Япония). Общее содержание липидов определяли по методу Блайя-Даера (Blight, Dayer, 1959).
Аминокислотный состав белка определяли на автоматическом аминокислотном анализаторе L-8800 (Hitachi, Япония). Подготовку проб осуществляли методом кислотного гидролиза биомассы водорослей (Баратова, Белякова, 1974; Остерман, 1985).
Жирнокислотный состав липидов определяли в виде метиловых эфиров жирных кислот (Carreau, Dubacq, 1978) на газожидкостном хроматографе GC-16А (Shimadzu, Япония), с пламенно-ионизационным детектором, капиллярной колонкой средней полярности (Carbowax-20) в токе гелия 28 см/с, температурой инжектора и детектора 240°С, термостата – 190°С, время анализа до 60 мин. Идентификацию жирных кислот проводили с помощью индексов удерживания жирных кислот, которые определялись индивидуально для каждой жирной кислоты.
Результаты и обсуждение
Содержание липидов в сахарине японской, добытой в прибрежных водах Приморского края, находится в диапазоне 1,0-1,7%, в водорослях из зал. Анива – 0,3-0,5%. Одной из основных идентификационных характеристик липидов водорослей является их жирнокислотный состав, который представлен в табл.1.
Таблица 1. Состав жирных кислот общих липидов сахарины японской побережья Приморского края (Японское море) и зал. Анива (Охотское море)
(% от суммы жирных кислот).
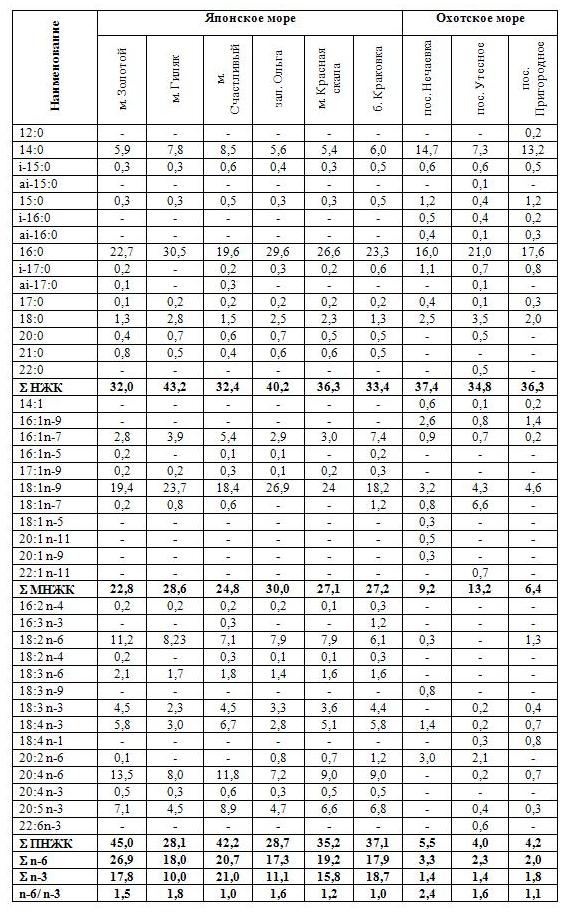
Жирнокислотный состав общих липидов исследованных водорослей Японского моря включает 26 жирных кислот (ЖК) с длиной цепей от 14 до 20 атомов (табл. 1). Доминирующими ЖК являются насыщенные (НЖК) и полиненасыщенные жирные кислоты (ПНЖК). Содержание ПНЖК в водорослях из исследованных районов находится в интервале от 28,1 до 45,0% и зависит от места произрастания. Наибольший уровень ПНЖК обнаружен в водорослях из акватории мысов Золотой и Счастливый – 45,0% и 42,2% соответственно (табл. 1). ПНЖК представлены диеновыми, триеновыми, тетраеновыми и пентаеновыми жирными кислотами. При этом на долю серии n-6 приходилось от 17,3 до 26,9%, серии n-3 – 11,1-21,01%. Основными ПНЖК являются арахидоновая (С20:4 n-6) и линолевая (С18:2 n-6) жирные кислоты, которые необходимы для нормальной жизнедеятельности организма человека и животных. Также в составе липидов исследованных водорослей Японского моря было выявлено 4,5-8,9% эйкозапентаеновой кислоты (С20:5 n-3), что сопоставимо с ее содержанием в треске, сельди и форели (Гладышев, 2012). Эйкозапентаеновая кислота является важным структурным компонентом клеточных мембран всех органов и тканей. Наибольшее содержание данной кислоты определено в водорослях из акватории м. Счастливый.
Удельный вес насыщенных ЖК в сахарине японской из исследованных районов побережья Приморского края варьирует от 32,0 до 43,2%. НЖК главным образом представлены пальмитиновой (С16:0) и миристиновой (С14:0) кислотами – 19,6-30,5% и 5,4-8,5%, соответственно (табл. 1).
Доля мононенасыщенных жирных кислот (МНЖК) составляет 22,8-30,0%. Максимальное их количество определено в водорослях из акватории б. Ольга. Из моноеновых жирных кислот преобладает олеиновая (С18:1n-9), на нее приходится до 26,9% от общего содержания липидов.
Жирнокислотный состав общих липидов сахарины японской Охотского моря отличается большим разнообразием ЖК. В их составе обнаружено 31 вид ЖК с длиной цепи от 12 до 22 атомов (табл. 1). Стоит отметить, что для исследованных водорослей Охотского моря характерно высокое содержание только НЖК (34,8-46,3%), при этом их количество сопоставимо с таковым в водорослях из Японского моря. В составе НЖК в следовых количествах были обнаружены лауриновая (С12:0) и бегеновая (С22:0) жирные кислоты, которые отсутствовали в качественном составе липидов водорослей Японского моря.
Уровень мононенасыщенных ЖК водорослей зал. Анива в среднем не превышает 9,6%, что в 2,8 раза ниже, чем у водорослей Японского моря. В составе липидов водорослей из зал. Анива были обнаружены С14:1, С16:1n-9 жирные кислоты. В водорослях из акватории пос. Нечаевка был выявлен ряд моноеновых кислот (С18:1n-5, С20:1n-11 и С20:1n-9), которые не были обнаружены в макрофитах из других районов Охотского и Японского морей.
Исследованные водоросли Охотского моря характеризуются низким содержанием полиненасыщенных кислот (менее 6%). Отличительной особенностью сахарины японской из прибрежных вод пос. Утесное является наличие молекул НЖК, МНЖК и ПНЖК с 22 атомами углерода – С22:0, С22:1n-1, C22:6n-3, которые не были обнаружены в водорослях из других районов Японского и Охотского морей. Стоит отметить, что докозагексаеновая кислота (C22:6n-3) играет важную роль в функционировании нервной системы (Гладышев, 2012), поэтому ее обнаружение в промысловых водорослях представляет большой интерес.
Анализируя состав жирных кислот водорослей из всех исследованных районов Японского и Охотского морей, можно констатировать, что большинство жирных кислот содержат четное число атомов углерода. На долю кислот С16 и С18 ряда, которые участвуют в образовании клеточных мембран (Гудвин, Мерсер, 1986; Napolitano, 1998), приходится от 60 до 80%. Среди насыщенных ЖК были отмечены молекулы с нечетным числом атомов – С15:0, С17:0, С21:0, которые редко встречаются в природе. В составе липидов исследованных водорослей определены длинноцепочечные молекулы ЖК (более 18 атомов углерода), что свойственно большинству гидробионтов и не характерно для наземных растений.
Установлено, что одной из основных причин, приводящих к вариациям в составе липидов сахарины японской, является район её произрастания. Так водоросли Японского моря отличаются значительным удельным весом моноеновых и полиеновых ЖК. Стоит также отметить, что водоросли Японского моря имеют высокий удельный вес n-3 ПНЖК (10-21%) по сравнению с макрофитами Охотского моря. Однако соотношение n-6 и n-3 ПНЖК в сахарине японской из обоих морей составляет 1,0:1 – 2,4:1, что соответствует рекомендациям Национального института здоровья США и японских национальных фондов (Davis, Kris-Etherton, 2003) и считается оптимальным для предотвращения сердечнососудистых заболеваний. А высокое содержание эйкозапентаеновой кислоты в водорослях Японского моря показывает высокую пищевую ценность и перспективность этого сырья для разработки диетических геродиетических продуктов. На основании проведенных исследований были отмечены районы с высоким содержанием насыщенных ЖК в сахарине японской – акватория м. Гиляк, мононенасыщенных – зал. Ольга, полиненасыщенных – м. Золотой, в том числе n-3 ПНЖК – м. Счастливый.
Общее количество белка в сахарине японской из прибрежных вод Приморского края варьировало от 4,0 до 8,4%, Охотского – от 7,0 до 9,9%. Белок исследованных водорослей состоит из 17 аминокислот, в том числе из 7 незаменимых (НАК) – валин, изолейцин, лейцин, лизин, метионин, треонин, фенилаланин и двух незаменимых для детей – аргинин и гистидин (табл. 2). Содержание триптофана в силу методических особенностей определено не было. Количество незаменимых аминокислот варьировало от 19,0 до 32,7%.
Известно, что биологическая ценность белка зависит от баланса его аминокислотного состава. Количество треонина и валина в белках большинства исследованных водорослей соответствует или незначительно превышает нормы, рекомендованные ФАО/ВОЗ. Валин является одним из главных компонентов в росте и синтезе тканей тела и вместе с лейцином служит источником энергии в мышечных клетках. По содержанию лейцина белки водорослей из акваторий м. Счастливый (Японское море) и пос. Нечаевка (Охотское море) превышают норму. Белки исследованных водорослей Японского и Охотского морей лимитированы по метионину. Данная аминокислота была определена только в сахарине японской из акватории м. Счастливый (Японское море). В целом, все незаменимые аминокислоты представлены в максимальном количестве в белках водорослей Охотского моря.
Таблица 2. Аминокислотный состав белка сахарины японской побережья Приморского края (Японское море) и зал. Анива (Охотское море) (г/ 100 г белка)
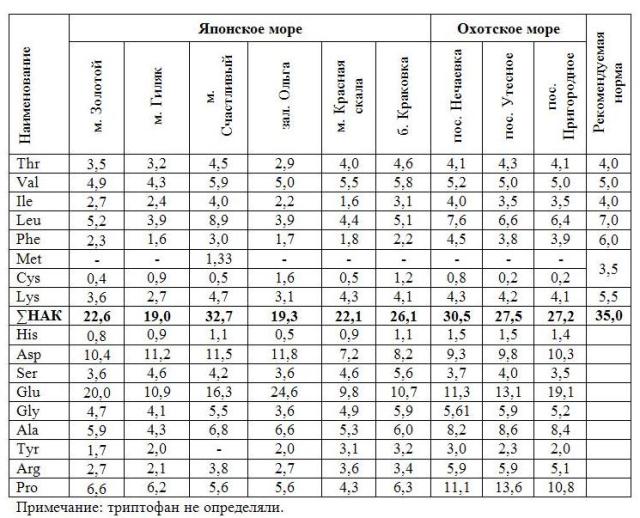
Что касается заменимых аминокислот, то независимо от места сбора водорослей в составе их белка преобладают дикарбоновые аминокислоты, а именно глутаминовая и аспарагиновая, составляющие 23-45% от суммы всех аминокислот. Данные кислоты имеют большое значение в обмене веществ в организме, участвуя в процессах торможения в нервной системе. Глутаминовая кислота является основной дикарбоновой кислотой исследуемых водорослей, её доля в водорослях Японского моря составляет 9,8-24,6%, Охотского – 11,3-19,1%. По количеству аспарагиновой кислоты водоросли исследуемых морей отличаются незначительно – 7,2-11,8%.
Для сахарины японской Охотского моря характерно высокое содержание пролина – в 2 раза выше, чем в водорослях Японского моря. Пролин отвечает за формирование коллагена в организме человека. В составе заменимых аминокислот макрофитов Охотского моря также определено в 1,5-2,0 раза больше аланина и аргенина соответственно.
Таким образом, приведенные в настоящей работе данные позволяют получить представление о различиях в длине и уровне насыщенности жирных кислот, а также о вариациях аминокислотного состава сахарины японской в зависимости от места ее произрастания. Липиды водорослей Японского моря отличаются значительным удельным весом моноеновых и полиеновых ЖК, в том числе n-3 серии. Для липидов сахарины японской Охотского моря характерно наличие большего числа наименований ЖК, которые отсутствовали в качественном составе липидов водорослей Японского моря, например лауриновая, бегеновая, докозагексаеновая.
Белки сахарины японской содержат практически весь комплекс незаменимых аминокислот, содержание которых выше в макрофитах Охотского моря. Среди заменимых аминокислот сахарины японской Охотского моря определено больше аланина, аргенина и пролина.
Список литературы
1. Баратова Л.А., Белякова Л.П. Определение аминокислотного состава белков // Методы биохимического эксперимента. М., 1974. С. 1-36.
2. Боголицин К.Г., Каплицин П.А., Ульяновский Н.В. Пронина О.А. Комплексное исследование химического состава бурых водорослей Белого моря// Химия растительного сырья. 2012. № 4. С. 153-160.
3. Воскобойников Г.М., Макаров М.В., Облучинская Е.Д., Рыжик И.В., Малавенда С.В. Макрофиты Баренцева моря: биологические особенности и перспективы использования // Формирование основ современной стратегии природопользования в Евро-Арктическом регионе. Апатиты: Изд-во КНЦ РАН, 2005. С. 316-328.
4. Герасименко Н.И., Бусарова Н.Г., Моисеенко О.П. Сезонные изменения в содержании липидов, жирных кислот и пиг ентов бурой водоросли Costaria coatata// Физиология растений. 2010. Т. 57. № 2. С. 217-2223.
5. Герасименко Н.И., Мартыяс Е.А., Бусарова Н.Г. Состав липидов, биологическая активность липидов и фотосинтетических пигментов водорослей семейств Laminariaceae и Alariaceae// Химия природных соединений. 2012. Т. № 5. С.662-666.
6. Гладышев М.И. Незаменимые полиненасыщенные жирные кислоты и их пищевые источники для человека// Journal of Siberian Federal University. Biology. 2012. V. 4. P. 352-386.
7. Гудвин Т., Мерсер Э. Введению в биохимию растений: в 2-х т.: пер. с англ. М., 1986. Т. 1. 393 с.
8. Евсеева Н.В., Репникова А.Р. Ресурсы промысловых водорослей Сахалино-Курильского региона// Рыбпром. 2010. № 3. С.14-21.
9. Остерман Л.А. Хроматография белков и нуклеиновых кислот: монография. М. , 1985. 536 с.
10. Состояние промысловых ресурсов. Прогноз общего вылова гидробионтов по Дальневосточному рыбохозяйственному бассейну на 2014 г. (краткая версия). – Владивосток: ТИНРО-Центр, 2014. – С. 298-310.
11. Blight E.G., Dayer W. J. A rapid method of total lipid extraction// Canad. J. Biochem. Phisiol. 1959. № 37. Р. 911-917.
12. Carreau J.P., Dubacq J.P. Adaptation of macro-scale method to the micro-scale for fatty acid methyl transesterification of biological lipid extracts// J. Chromatogr. 1978. V. 151. № 3. P. 384-390.
13. Carroll D.N., Roth M.T. Evidence for the Cardioprotective Effects of Omega-3 Fatty Acids// Ann. Pharmacotherapy. 2002. V. 36. P.1950-1956.
14. Davis B.C, Kris-Etherton P.M. Achieving optimal essential fatty acid status in vegetarians: current knowledge and practical implications// American Journal of Clinical Nutrition. 2003. V. 78. Р. 640-646 .
15. Napolitano G.T. Fatty acids as trophic and chemical markers in freshwater ecosystems// Lipids in freshwater ecosystems/ Eds. Arts M.T., Wainman B.C. New York. – 1998. – P. 21-25.
16. Honya M., Kinoshita T., Ishikawa M., Mori H., Nisizawa K. Seasonal Variation in the Lipid Content of Cultured Laminaria japonica: Fatty Acids, Sterols, b-Carotene and Tocopherol// J. Appl. Phycol. 1994. V. 6. P. 25-29.
17. Hou X., Yan X., Chai C. Chemical species of iodine in some seaweeds. II. Iodine-bound biological macromolecules // J. of Radioanalytical and Nuclear Chemistry. 2000. V. 245. № 3. P. 461-467.
18. Kim M.K., Dubacq J.P., Thomas J.C., Giraud G. Seasonal Variation of Triacylglycerols and Fatty Acids in Fucus serratus // Phytochemistry. 1996. V. 43. P. 49-55.
19. Sanina N.M., Goncharova S.N., Kostetsky E.Y. Fatty acid composition of individual polar lipid classes from marine macrophytes// Phytochemistry. 2004. V. 65. P. 721-730.
20. Sanina N.M., Goncharova S.N., Kostetsky E.Y. Seasonal Changes of Fatty Acid Composition and Thermotropic Behavior of Polar Lipids from Marine Macrophytes// Phytochemistry. 2008. V. 69. P. 1517-1527.
опубликовано - декабрь 2014 г.
Features of amino acid and fatty acid compositions of Saccharina japonica Okhotsk and Japan seas
Olga N. Guruleva, Elena L. Koneva, Tatiana I. Vishnevskaya, Irina A. Kadnikova,
Konstantine G. Pavel', Natalia M. Aminina
The main fatty acids of general lipids contained in Saccharina japonica from the Japan and the Okhotsk Seas are the C16:0, C18:1n-9 fatty acids. The fatty acids С18:2n-6, С20:4n-6 dominate in the Japan Sea algae and the fatty acid С14:0 prevails in Saccharina japonica from the Okhotsk Sea. In the Japan Sea algae, it’s determined up to 8.9% of eicosapentaenoic acid (C20:5n-3). The share of polyunsaturated fatty acids (PUFAs) of the n-3 series varies from 1.4 % to 21.0 %, the n-6 series from 2.0% to 26.9% depending on the algae gathering area. The n-6/n-3 ratio of polyunsaturated fatty acids was 1.0:1 – 2.4:1. Based on this research, the habitat areas of Saccharina japonica with high content of saturated fat, monounsaturated fat, polyunsaturated fat including the n-3 series are defined.
17 amino acids are defined in the protein of investigated algae, including 7 irreplaceable amino acids: valine, isoleucine, leucine, lysine, methionine, threonine, phenylalanine. Noted that the content of irreplaceable amino acids in the Okhotsk Sea algae higher than in the Japan Sea saccharine. Aspartic,glutamic acids and praline predominate among the nonessential amino acids.
Keywords: brown seaweed; Saccharina japonica; fatty acids; amino acids.
Об авторах
Гурулева Ольга Николаевна – Guruleva Olga N.
кандидат технических наук
научный сотрудник Лаборатории проблем рационального использования водорослей ФГУП ТИНРО-Центр (FSUI «Pacific Scientific Research Fisheries Centre» – «TINRO-centre»)
guruleva@tinro.ru
Конева Елена Леонидовна - Koneva Elena L.
кандидат технических наук
научный сотрудник Лаборатории проблем рационального использования водорослей ФГУП ТИНРО-Центр (FSUI «Pacific Scientific Research Fisheries Centre» – «TINRO-centre»)
koneva@tinro.ru
Вишневская Татьяна Ивановна - Vishnevskaya Tatiana I.
кандидат технических наук
старший научный сотрудник Лаборатории проблем рационального использования водорослей ФГУП ТИНРО-Центр (FSUI «Pacific Scientific Research Fisheries Centre» – «TINRO-centre»)
vishnevskaya@tinro.ru
Кадникова Ирина Арнольдовна - Kadnikova Irina A.
доктор технических наук
ведущий научный сотрудник Лаборатории проблем рационального использования водорослей ФГУП ТИНРО-Центр (FSUI «Pacific Scientific Research Fisheries Centre» – «TINRO-centre»)
kadnikova@tinro.ru
Павель Константин Геннадьевич - Pavel' Konstantine G.
кандидат химических наук
старший научный сотрудник Лаборатории проблем рационального использования гидробионтов ФГУП ТИНРО-Центр (FSUI «Pacific Scientific Research Fisheries Centre» – «TINRO-centre»)
pavel@tinro.ru
Аминина Наталья Михайловна - Aminina Natalia M.
кандидат биологических наук
заведующая Лабораторией проблем рационального использования водорослей ФГУП ТИНРО-Центр (FSUI «Pacific Scientific Research Fisheries Centre» – «TINRO-centre»)
aminina@tinro.ru
Корреспондентский адрес: Россия, 690091, г. Владивосток, пер. Шевченко, 4, ФГУП ТИНРО-центр; телефон (423) 240-13-60
ССЫЛКА НА СТАТЬЮ:
Гурулёва О.Н., Конева Е.Л., Вишневская Т.И., Кадникова И.А., Павель К.Г., Аминина Н.М. Особенности аминокислотного и жирнокислотного составов Saccharina japonica Охотского и Японского морей // Вопросы современной альгологии. 2014. № 2 (6). URL: http://algology.ru/599
Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно.
Адрес - info@algology.ru
При перепечатке ссылка на сайт обязательна
К другим статьям Международной конференции «Экологическая физиология водных фототрофов: распространение, запасы, химический состав и использование». III Сабининские чтения.
К разделу СТАТЬИ.
Карта сайта

|
