|
|
|
Влияние различных источников азота на рост динофитовых водорослей Черного моря
Мансурова И. М., Стельмах Л. В. Irina M. Mansurova, Lyudmyla V. Stelmakh
Институт биологии южных морей им. А.О. Ковалевского (г. Севастополь)
УДК 582.252:577.113.3 (262.5)
Исследовано влияние нитратов, аммония и мочевины на рост 4 видов черноморских динофитовых водорослей в условиях низкой (10; 19 мкЭ×м-2×с-1) и высокой (95; 105 мкЭ×м-2×с-1) интенсивности света. Показано, что только у Prorocentrum cordatum не наблюдается различий по росту на всех трех формах азота. Для большинства видов выявлена видоспецифичная реакция на различные источники азотного питания. При высокой интенсивности света у Gyrodinium fissum удельная скорость роста на мочевине снижалась в 1,5 раза по сравнению с ростом на нитратах и аммонии. При низкой интенсивности света у Heterocapsa triquetra и P. pusillum увеличивалась продолжительность лаг-фазы на аммонии, тогда как на нитратах и мочевине она была в 2-3 раза короче. Ключевые слова: динофитовые водоросли; нитраты; аммоний; мочевина; удельная скорость роста.
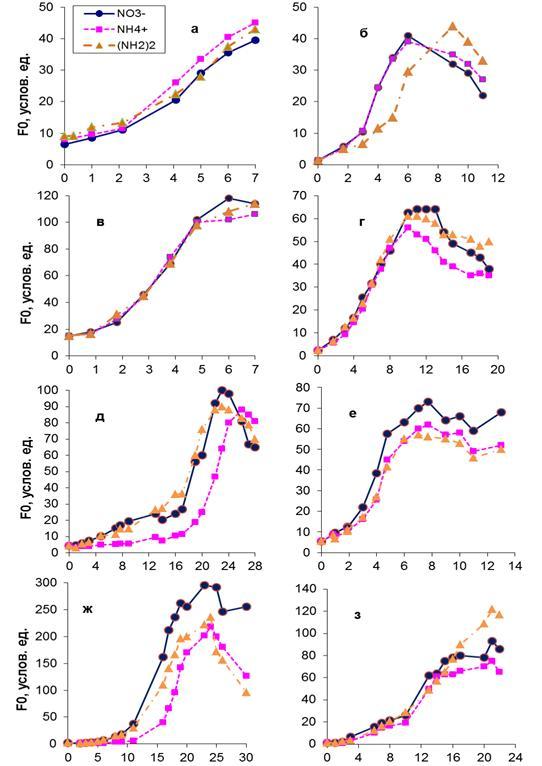
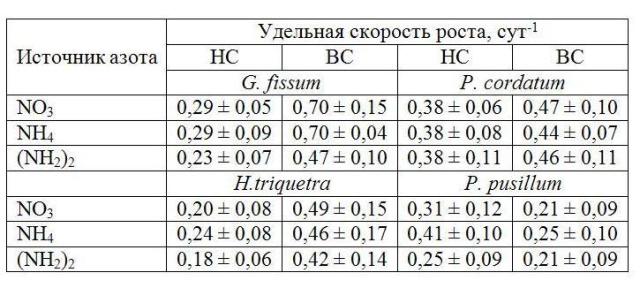
Известно, что в Черном море основную численность и биомассу фитопланктона, как правило, создают диатомовые (Bacillariophyta) и динофитовые (Dinophyta) водоросли. Соотношение между этими таксономическими группами претерпевает регулярную временную и пространственную изменчивость, которая обусловлена совместным влиянием абиотических факторов и биотическими взаимодействиями в планктоне. При изменении параметров среды, таких как свет, температура, количество питательных веществ, происходит перестройка таксономической и размерной структуры фитопланктона, изменяется количество видов внутри каждого таксона. Среди биогенных веществ, влияющих на развитие водорослей в море, важную роль играют соединения азота. Основными формами азота, присутствующими в морской воде, являются нитраты, аммоний и мочевина (Парсонс, 1982). Показано, что в одних случаях рост водорослей в культурах одинаково успешно осуществлялся на всех трех формах азота, в других – наблюдались различия (Dortch,1990; Levasseur et al, 1993; Lourenco et al., 2002). Основные результаты, представленные в литературе, были получены, как правило, для диатомовых видов водорослей, зеленых и цианобактерий (Dortch, 1990; Lourenco et al., 2002). Что касается динофитовых водорослей, то таких данных недостаточно для, того чтобы судить об особенностях роста этих видов на разных источниках азотного питания. Цель настоящей работы заключалась в исследовании влияния нитратов, аммония и мочевины на рост черноморских видов динофитовых водорослей в условиях культур. Материалы и методика Экспериментальные исследования были выполнены на альгологически чистых культурах динофитовых водорослей Gyrodinium fissum Levander, Prorocentrum cordatum Ostenfeld, Heterocapsa triquetra Ehrenberg и P. pusillum Schiller, представленных в коллекции Отдела экологической физиологии водорослей ИнБЮМ (г. Севастополь). В качестве питательной основы использовали раствор Гольдберга (Финенко и др., 2008). В трех вариантах среды использовали разные формы азота: нитраты, аммоний в конечной концентрации 100 мкМ и мочевину (50 мкМ). Водоросли экспонировали при двух интенсивностях света: низкой (10 и 19 мкЭ×м-2×с-1) и высокой (95 и 105 мкЭ×м-2×с-1). Адаптация к высокой интенсивности света осуществлялась в течение 1-х суток, а к низкой достигала 15 суток. Одновременно водоросли доводились до дефицита азота в среде, критерием которого были низкие значения относительной переменной флуоресценции хлорофилла in vivo (Fv/Fm), составлявшие ≤0,30 (Запара и др., 2008). После этого в среду, где росли водоросли, вносили одну из указанных форм азота. Температура в экспериментах составляла 20-22ºС. Прирост водорослей в накопительных культурах оценивали по величине переменной флуоресценции (F0), измеренной in vivo при открытых реакционных центрах фотосистемы II, которая изменяется пропорционально концентрации хлорофилла а в клетках водорослей. Интенсивность света измерялась с помощью люксметра Ю-116. Для перехода от освещенности в люксах к энергетическим единицам использовался коэффициент перехода 1000 лк = 17,2 мкЭ×м-2×с-1 (Парсонс, 1982). Результаты и обсуждение Выполненные исследования показали, что только для одного вида водорослей – P. cordatum – различия по росту (по величине F0) на всех трех формах азота не наблюдались как при низкой, так и при высокой интенсивности света (рис. 1 в, г). При этом средние значения удельной скорости роста в экспоненциальной фазе на разных формах азота были близки (табл. 1). Подобные исследования, проведенные ранее с этим видом при более высокой концентрации соединений азота (1,18 мкМ), температуре 20-23°С и высокой интенсивности света (300 мкЭ×м-2×с-1) в цикле свет:темнота (12:12 ч), также не показали различий в скорости роста на нитратах и мочевине (Lourenco et al., 2002). Тогда как на аммонии культура не росла, так как столь высокие концентрации являются, по мнению авторов, токсичными. В условиях слабого света (10, 19 мкЭ×м-2×с-1) рост H. triquetra на нитратах и мочевине уже в начальный период развития накопительной культуры (1–7 сутки) был существенно выше (в 3–4 раза), чем на аммонийном азоте (рис. 1, д). Подобные результаты получены и для P. pusillum (рис. 1, ж). Причина таких различий для данных видов водорослей связана с тем, что лаг-фаза их развития на аммонии была более продолжительна и составляла 14 и 11 суток соответственно, что приблизительно в 2–3 раза больше, чем на нитратах и мочевине. При насыщающей рост интенсивности света лаг-фаза у H. triquetra и P. pusillum не проявлялась, а средние значения удельной скорости роста достоверно не различались. В результате на всех трех формах азота прирост был одинаков. Для G. fissum в оптимальных для роста световых условиях (105 мкЭ×м-2×с-1) прирост (по F0) в накопительной культуре на мочевине был ниже, чем на нитратах и аммонии (рис. 1, б). Удельная скорость роста на данной форме азота составила в среднем 0,47 сутки-1, тогда как на других источниках азота она была почти в 1,5 раза выше. Подобные результаты были получены ранее для динофитовой водоросли Gymnodinium sanguineum (Levasseur et al, 1993). Показано, что при 18°С и таких же исходных концентрациях азота, как и в наших экспериментах, значения удельной скорости роста для этого вида при насыщающей рост интенсивности света на нитратном и аммонийном азоте существенно не отличались, а на мочевине были в 2,3-2,4 раза ниже. По нашим данным, на низком свету рост G. fissum на всех трех формах азота был одинаков.
Рис. 1. Рост динофитовых водорослей в накопительных культурах на нитратах, аммонии и мочевине (F0 – величина переменной флуоресценции в условных единицах): Fig. 1.Growth of dinoflagellates in batch cultures on nitrates, ammonium and urea (F0 – fluorescence in conventional units):
Таблица 1. Удельная скорость роста водорослей на различных источниках азота в экспоненциальной фазе роста на низком (НС= 10; 19 мкЭ×м-2×с-1) и высоком свету (ВС= 95; 105 мкЭ×м-2×с-1). Table 1. Specific growth rate of microalgae on different nitrogen sources in exponential growth at low (LL = 10; 19 µmol×m-2×s-1) and high light (HL = 95; 105 µmol×m-2×s-1).
Таким образом, для большинства исследованных видов динофитовых водорослей выявлены различия по росту на различных источниках азота. Эти различия наблюдались при низких и высоких интенсивностях света. Выводы 1. В условиях слабого освещения у двух видов динофитовых водорослей H. triquetra и P. pusillum прирост в накопительной культуре на начальном этапе роста на аммонийном азоте был в 2 – 4 раза ниже, чем на нитратах и мочевине. В оптимальных для роста световых условиях у G. fissum в середине экспоненциальной фазы роста прирост на мочевине был в 1,5 раза ниже, чем на нитратах и аммонии. Таким образом, можно говорить о видоспецифичной реакции большинства исследованных видов динофитовых водорослей на различные источники азота, которая проявлялась либо при низких, либо при высоких интенсивностях света. 2. Для одного вида – P. cordatum – не наблюдалось различий по росту на трех источниках азота ни в лаг-, ни в экспоненциальной фазе роста как на низком, так и на высоком свету. Следовательно, данный вид наиболее конкурентоспособен в борьбе за азот среди исследованных динофитовых водорослей.
Авторы выражают искреннюю благодарность научному сотруднику отдела экологической физиологии водорослей ИнБЮМ (г. Севастополь) Акимову А.И. за консультации и помощь в планировании экспериментальной работы.
Список литературы 1. Запара Е. В., Белевич Т. А., Ильяш, Л. В. Конкурентные параметры планктонных водорослей Белого моря при эксплуатации органического и минерального азота // Бюллетень Московского общества испытателей природы. Отдел Биологический. 2008. Т. 113, №3. С. 43-49. 2. Парсонс Т. Р., Такахаши М., Харгрейв В. Биологическая океанография. М.: Легкая и пищевая промышленность,1982. С. 89-91. 3. Финенко З. З., Стельмах Л. В., Галатонова О. А., Бабич И. И. Культивирование водорослей в лабораторных условиях / Токарев Ю. Н., Финенко З. З., Шадрин Н. В. Микроводоросли Черного моря: проблемы сохранения биоразнообразия и биотехнологического использования. Севастополь: Экоси-Гидрофизика, 2008. С. 186- 200. 4. Dortch, Q. 1990. The interaction between ammonium and nitrate uptake in phytoplankton // Marine Ecology Progress Series. 1990. Vol. 61. P. 183-201. 5. Levasseur, M., Thompson, P. A,. Harrison, P. J. Physiological acclimation of marine phytoplankton to different nitrogen sources // J. Phycol. 1993. Vol. 29. P. 587–595. 6. Lourenco, S.O., Barbarino, E., Mancini-Filho, J., Schinke, K.P., Aidar, E. Effects of different nitrogen sources on the growth and biochemical profile of 10 marine microalgae in batch culture: an evaluation for aquaculture // Phycologia. 3002. Vol. 41, №2. P. 158–168. опубликовано - ноябрь 2014 г.
Effect of the different nitrogen sources on the growth of Black Sea dinoflagellates Irina M. Mansurova, Lyudmyla V. Stelmakh Effect of the nitrogen, ammonium and urea on the growth of four Black Sea dinoflagellates species was investigated at low (10, 19 µmol×m-2×s-1) and hight light (95, 105 µmol×m-2×s-1). It was found that there were no differences in growth in all three forms of nitrogen only for P. cordatum. Species-specific responses to different sources of nitrogen nutrition were observed for most species. It was expressed in decrease of the specific growth rate on urea in 1,5 times compared with growth on nitrate and ammonium for G. fissum at high light. At low light intensity the lag phase for H. triquetra and P. pusillum was in 2 - 3 times longer on ammonium whereas on nitrate and urea. Keywords: dinoflagellates; nitrates; ammonia; urea; specific growth rate.
Об авторах Мансурова Ирина Мяулитовна – Mansurova Irina Maylitovna ведущий инженер Института биологии южных морей им. А.О. Ковалевского, г. Севастополь (A.O. Kovalevsky Institute of Biology of Southern Seas, Sevastopol) ira.mansurova2013@yandex.ua Стельмах Людмила Васильевна – Stelmakh Lyudmyla Vasilievna кандидат биологических наук lustelm@mail.ru Корреспондентский адрес: 299011 г. Севастополь, пр. Нахимова, 2, Институт биологии южных морей им. А.О. Ковалевского
ССЫЛКА НА СТАТЬЮ: Мансурова И. М., Стельмах Л. В. Влияние различных источников азота на рост динофитовых водорослей Черного моря // Вопросы современной альгологии. 2014. № 2 (6). URL: http://algology.ru/598
При перепечатке ссылка на сайт обязательна
На ГЛАВНУЮ
|
|||
|
| ||