из Материалов Международной конференции «Экологическая физиология водных фототрофов: распространение, запасы, химический состав и использование» III Сабининские чтения
29 ноября - 29 декабря 2014 г.
Экология диатомовых водорослей перифитона особо охраняемых природных территорий на примере Дальневосточного морского государственного природного биосферного заповедника
(залив Посьета, Японское море)
Ecology of periphyton diatoms of specially protected natural territories for example to Far-Eastern Marine State Natural Biosphere Reserve
(Posyet Bay, Japan Sea)
Бегун А.А.
Andrey A. Begun
Институт биологии моря им. А.В. Жирмунского ДВО РАН
УДК 579.26+582.26+58.009+574.587
Исследованы видовой состав и экология диатомовых водорослей перифитона навигационных буев в акваториях заливе Посьета Японского моря. Показано, что для некоторых акваторий залива Посьета характерно доминирование индикаторов значительного органического загрязнения на фоне низких показателей видового богатства диатомового перифитона. Особенности видового состава и экологии диатомового перифитона в значительной степени определяются уровнем антропогенного загрязнения акваторий, прилегающих к Морскому заповеднику.
Ключевые слова: перифитон; диатомовые водоросли; сапробность; навигационные буи; Морской заповедник; залив Посьета; Японское море.
Введение
На юге Дальнего Востока Российской Федерации расположен редкий по красоте уголок живой природы, где тайга встречается с океаном, а холодные воды с тропическими, и где сложились уникальные природные комплексы растений и животных, аналогов которым нет больше нигде в мире. В этом удивительном регионе расположен Дальневосточный морской государственный природный биосферный заповедник (далее – Морской заповедник), созданный в 1978 г. Он является единственным в нашей стране настоящим морским заповедником, свыше 98% площади которого – это акватория. По мнению специалистов, для успешного функционирования Морского заповедника, как природоохранной зоны федерального значения и объекта морского природного наследия, крайне важен мониторинг его биоразнообразия, при котором на основании ранее проведенной инвентаризации биоты и ее количественного распределения дается экологическая оценка ее нынешнего состояния (Малютин, Гульбина, 2012). Необходимость исследований биоты в акваториях, подверженных влиянию судового балласта, к которым относятся порты зал. Посьета, во многом связана с началом проведения Институтом биологии моря ДВО РАН оценок экологических рисков и мониторинга видов-вселенцев на шельфе Дальневосточных морей России (Звягинцев и др., 2012).
Проводимые ранее исследования диатомовых водорослей микрофитобентоса носили единовременный характер и были сделаны в нескольких локальных точках, прилегающих к западному участку Морского заповедника – в бух. Рейд Паллада и Троицы и к восточному участку – о-ву Большой Пелис (архипелаг Римского Корсакова) (Николаев, 1976; Рябушко, 1986). Из альгофлоры здесь основное внимание уделялось морскому фитопланктону (260 видов), пресноводным водорослям (1032), а также макрофитам (170), в то время как флора морского микрофитобентоса зал. Посьета не вошла в сводку биоты Морского заповедника, а диатомовые водоросли перифитона на искусственных субстратах вообще не были исследованы.
Микроводоросли перифитона являются важнейшим автотрофным компонентом любых водных экосистем как первичные продуценты органического вещества и начальное звено биопродукционных процессов. Диатомовые водоросли, составляющие основу фитоперифитона, имеют малые размеры и короткий жизненный цикл, что позволяет им быстро реагировать на изменение экологического состояния в биотопе, а по систематическим, количественным и морфологическим показателям и сапробности их ведущих видов судят о состоянии «здоровья» морской среды (Оксиюк, Давыдов, 2006; Бегун и др., 2010; Бегун, 2012). Для изучения экологии диатомового перифитона на территории Морского заповедника в качестве наиболее удобных модельных объектов мы рассматриваем буи плавучего навигационного ограждения. Они представляют собой экспонируемый в море на определенной глубине в фиксированный временной интервал субстрат, подверженный формированию макрообрастания как растительного, так и животного происхождения, который, в свою очередь, служит субстратом для оседания и развития диатомового перифитона.
Цель настоящей работы заключалась в изучении видового состава и экологии диатомовых водорослей перифитона на буях плавучего навигационного ограждения в акваториях зал. Посьета Японского моря, прилегающих к Морскому заповеднику и в различной степени подверженных антропогенной нагрузке.
Материалы и методы
Материалом для исследования послужили качественные пробы микроводорослей с 10 буев плавучего навигационного ограждения, выставленные в апреле 2011 г. перед открытием навигации маломерного флота, в акваториях зал. Посьета Японского моря, прилегающих к западному и восточному участкам Морского заповедника на глубине 0,5–2,0 м от поверхности воды, демонтированные в ноябре 2011 г. (рис. 1). Отбор материала произведен 22 ноября на борту гидрографического судна тихоокеанского флота "ГС-84". Приводим описание координат исследуемых навигационных буев: № 1 (420 37,801 N 130047,950 Е) и 4 (420 38,04 N 130047,95 Е) располагался в районе косы Назимова, отделяющей бух. Рейд Паллада от бух. Экспедиции, № 2 (420 37,94 N 130048,04 Е) – около о. Черкасского, № 3 (420 38,82 N 130048,78 Е) – в районе рифа бух. Новгородской, № 5 (420 38,70 N 130048,053 Е) – в районе м. Рязанова, разделяющего бух. Портовую и Порт-Посьет, № 6 (420 33,82 N130054,89 Е) – в районе банки Клыкова перед входом в бух. Рейд Паллада, №№ 7-9 (420 38,1 N 131005,7 Е, 420 38,5 N 131005,4 Е и 420 38,325 N 131005,13 Е, соответственно) – в районе о. Браузера (бух. Троицы) и № 10 (420 38,6 N 131005,1 Е) – в районе м. Шульца (вход в бух. Витязь) (Лоция…, 1984). Буи №№ 1–6 и 10 располагались в бухтах Рейд Паллада, Экспедиции, Новгородская, Портовая, Порт-Посьет, Витязь, не испытывающих влияния неочищенных сточных вод и иных источников загрязнения, и рассматривались в качестве фоновых, а буи №№ 7–9 находились в бух. Троицы, испытывающей хроническое антропогенное загрязнение со стороны морского порта «Зарубино» и рыбокомбината (Белан, 2001; Кобзарь, Христофорова, 2012).
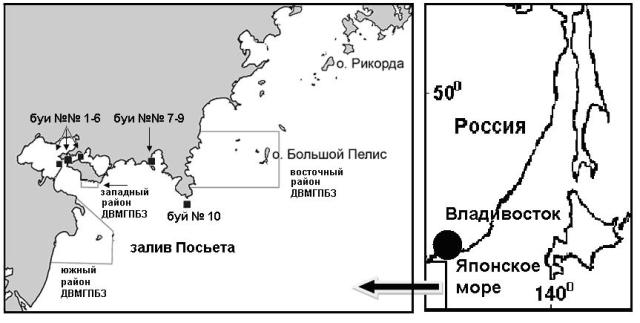
Рис. 1. Карта-схема района и станций исследования буев плавучего навигационного ограждения в акваториях зал. Посьета Японского моря, прилегающих к Морскому заповеднику, ноябрь 2011 г.
Пробы перифитона с поверхности навигационных буев отбирали специальным скребком-сачком с режущим краем длиной 10 см и острым углом наклона к исследуемой поверхности общей площадью 50х50 мм2. С каждого из исследуемых буев (рис. 2) с помощью синтетической щетки отбирали по 4 пробы перифитона: с поверхности зеленых водорослей-макрофитов, обрастающих верхнюю часть буя, со средней части самой бочки буя и ее трубы, и с якорной цепи. Полученные суспензии со смывами микроводорослей пропускали через капроновые фильтры «Nitex» («Sefar», Швейцария) с диаметром пор 20 и 80 микрон, соответственно. Фракции этих смывов концентрировали методом осаждения до 4–5 мл. Пробы фиксировали 4%-ным раствором формалина и Утермёлем до бледно-желтого цвета.

Рис. 2. Общий вид обросших буев плавучего навигационного ограждения в акваториях, прилегающих к Морскому заповеднику, в период их демонтажа в ноябре 2011 г.
Исследования постоянных препаратов проводили на световом микроскопе («Olympus BX41», объектив UPLanF1100х/1/.30) с использованием иммерсии. Для детальной идентификации диатомей приготовляли постоянные препараты по общепринятой методике (Диатомовые…, 1974). Образцы для сканирующей электронной микроскопии подготавливали по стандартной методике, включающей промывание клеток дистиллированной водой, их дегидратацию через серию спиртов (25, 50, 75, 96 и 100 %) и сушку на воздухе (Truby, 1997). На поверхность образцов в вакуумном посту «Edwards, AUTO 306» (Великобритания) наносили электропроводящее покрытие из золота. Препараты изучали под сканирующим электронным микроскопом «Leo-430» (Великобритания).
Результаты и обсуждение
В перифитоне навигационных буев, исследованных в акваториях зал. Посьета Японского моря, прилегающих к Морскому заповеднику, зарегистрировано 93 вида и внутривидовых таксонов (ввт) диатомовых водорослей, принадлежащих к классам Bacillariophyceae (59 % от общего числа видов), Coscinodiscophyceae (25 %) и Fragilariophyceae (16 %).
В акватории зал. Посьета впервые зарегистрировано 43 вида и ввт Bacillariophyta:
Achnanthes brevipes var. intermedia (Kütz.) Cleve,
Biddulphia arctica f. balaena (Ehrenb.) E.H. Jörgens,
B. biddulphiana (J.E. Smith) Boyer,
Campylodiscus fastuosus Ehrenb.,
Coscinodiscus granii Gough,
C. oculus-iridis Ehrenb.,
Cyclotella choctawhatcheeana Prasad,
Diatoma tenuis C. Agardh,
Dactyliosolen fragilissimus (Bergon) Hasle,
Dimeregramma minor (Greg.) Ralfs ex Pritch.,
Diploneis interrupta (Kütz.) Cleve,
Falcula media Voigt var. subsalina Proschk.-Lavr.,
Fragilaria capucina Desm.,
F. crotonensis Kitton,
Gyrosigma tenuissimum (W. Sm.) Grif. et Henfr.,
Halamphora cymbifera (Greg.) Levkov,
Guinardia striata (Stolterfoth) Hasle,
Hantzschia amphioxys (Ehrenb.) Grun.,
Haslea subagnita (Proschk.-Lavr.) I.V. Makar. et N.I. Kar.,
Lyrella clavata (Greg.)., D.G. Mann,
Melosira moniliformis var. subglobosa (Grun.) Hustedt,
Navicula cancellata var. retusa (Bréb.) Cleve,
N. cryptocephala Kütz.,
Neosynedra provincialis (Grun.) Williams et Round,
Neohuttonia reichardtii (Grun.) Hustedt,
Nitzschia acicularis (Kütz.) W. Sm.,
N. hybrida f. hyalina Proschk.-Lavr.,
N. palea (Kütz.) W. Sm.,
N. scapelliformis (Grun.) Grun.,
N. spathulata W. Sm.,
N. vermicularis (Kütz.) Hantzsch ex Rabenh.,
Odontella granulata (Roper) R. Ross,
Parlibellus rhombicus (Greg.) E.J. Cox,
Plagiogrammopsis vanheurckii (Grun.) Hasle, von Stosch & Syvertsen,
Plagiotropis elegans (W. Sm.) Grun.,
Pleurosigma intermedium W. Sm.,
P. naviculaceum Bréb.,
Rhizosolenia setigera Brightw.,
Synedra toxoneides Castracane,
Tabellaria fenestrata (Lyng.) Kütz.,
Thalassiophysa hyalina (Grev.) Paddock et P.A. Sims,
Tryblionella hungarica (Grun.) D.G. Mann,
Undatella lineolata (Ehrenb.) L.I. Ryab.
Ранее в микрофитобентосе различных типов субстратов (макрофиты, каменистые и рыхлые грунты) зал. Посьета зарегистрировано 295 видов диатомовых водорослей (Николаев, 1976), однако после ревизии списка этих видов их количество составляет около 280 (Рябушко 1986, 2013). Сравнение видов, обнаруженных в перифитоне навигационных буев, с данными по микрофитобентосу природных субстратов залива (Николаев, 1976; Рябушко, 1986), показало, что около 30 видов являются общими.
Экологический анализ флоры диатомовых водорослей показал, что все найденные виды представлены подвижными (52,4%), прикрепленными (31,3%) и прикреплено-подвижными (13,7%) формами, которые, в свою очередь, подразделяются на одиночноживущие (53%) и колониальные (47%). По отношению к биотопу 60% всех видов являются бентосными, 27% – бентопланктонными (тихопелагическими) и 13% – фитопланктонными (рис. 3). По отношению к солености преобладают эвригалинные виды, из них морские составляют 58%, морские и солоноватоводные – 20%; в незначительной степени представлены солоноватоводные (7%), пресноводно-солоноватоводные (9%) и пресноводные (6%). По отношению к pH среды доминируют алкалифильные виды (98%). Из всех найденных видов к типичным эпифитам и обрастателям твердых субстратов относится около 20 видов диатомовых водорослей, другая часть видов принадлежит к флоре песчаных и илистых грунтов (эпипсаммон и эпипелон), попавших на субстрат в период взмучивания вод, либо к формам, осевшим из толщи воды. Анализ фитогеографии диатомовых водорослей, тесно связанной с их экологией, показал, что из всех встреченных видов 42% являются космополитами, 26% – бореальными и 14% – аркто-бореально-тропическими видами.
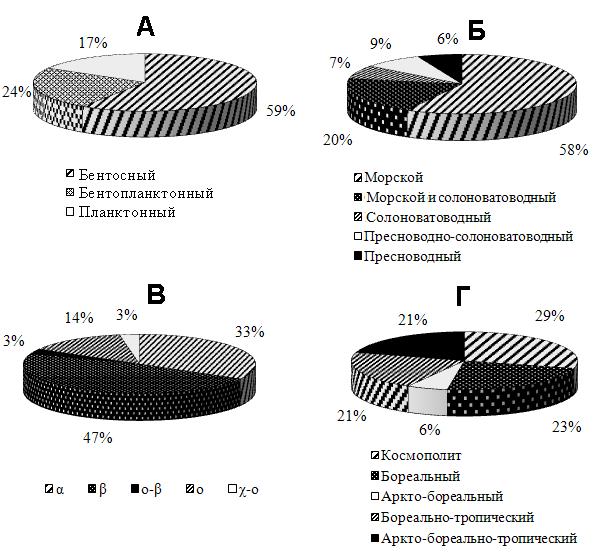
Рис. 3. Эколого-фитогеографическая характеристика диатомовых водорослей перифитона на навигационных буях в акваториях, прилегающих к Морскому заповеднику, в ноябре 2011 г.
А – жизненная форма, Б – отношение видов к солености, В – сапробиологическая характеристика
(α – альфа-мезосапробионт, β – бета-мезосапробионт, ο-β – олиго-бета-мезосапробионт,
ο – олигосапробионт, χ-ο – ксено-олигосапробионт), Г – эколого-фитогеографическая принадлежность.
Эколого-фитогеографическая структура диатомовых водорослей перифитона акваторий зал. Посьета, прилегающих к Морскому заповеднику, достаточно широка – от морских форм, обитающих на литорали и сублиторали морей с морской соленостью, до соловатоводных и пресноводных, попадающих в пелагиаль прибрежной зоны морей в результате влияния берегового стока. В частности, виды Tabularia fasciculata (Agardh) Williams et Round, Achnanthes brevipes var. intermedia (Kütz.) Cleve, Parlibellus delognei (V.H.) E.J. Cox, Gyrosigma tenuissimum (W. Sm.) Grif. et Henfr., Melosira lineata (Dillw.) Agardh, M. moniliformis (Grun.) Hustedt и Halamphora cymbifera (Greg.) Levkov (рис. 4) являются морскими и солоноватоводными, эвритермными и эвригалинными видами, широко распространенными как в морях, так и в эстуариях умеренной зоны и относящимися к бореальному, аркто-бореальному и космополитическому фитогеографическим комплексам (Гусляков и др., 1992; Герасимюк и др., 2011; Рябушко, 2013).
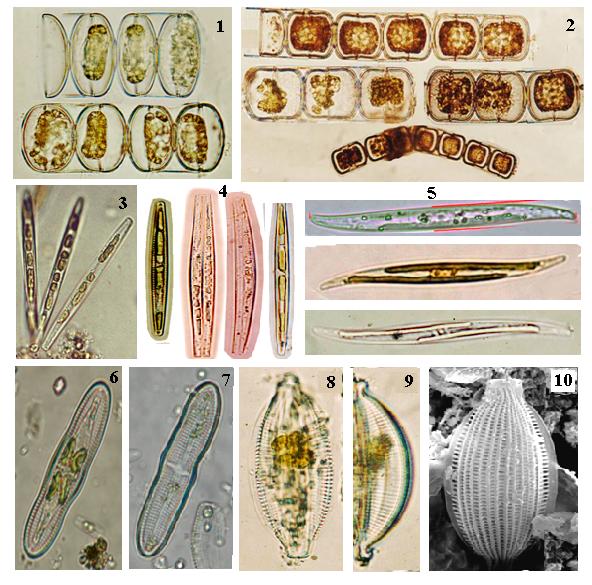
Рис. 4. Виды диатомовых водорослей перифитона навигационных буев в акваториях зал. Посьета Японского моря, прилегающих к Морскому заповеднику, ноябрь, 2011 г.
1 – Melosira moniliformis, 2 – M. lineata, 3, 4 – Tabularia fasciculata, 5 – Gyrosigma tenuissimum, 6, 7 – Achnanthes brevipes var. intermedia, 8–10 – Halamphora cymbifera (1-9 – световая микроскопия, 10 – сканирующая электронная микроскопия).
Некоторые виды диатомовых водорослей, указанные в зал. Посьета, являются эвритермными, встречающиеся круглогодично независимо от изменений температуры воды, которая подвержена значительным изменениям в течение года за счет конвективного перемешивания вод и штормов. Фитогеография диатомовых водорослей в акваториях, прилегающих к Морскому заповеднику, во многом связана с факторами течений и циркуляцией вод, оказывающих существенное влияние на проникновение тепловодной фауны и флоры в заливы и бухты Приморья. Как было показано ранее, диатомовые водоросли северо-западной части Японского моря принадлежат в основном к широко-бореальному комплексу с аркто-бореальными и космополитическими элементами флоры (Николаев, 1976; Рябушко, 1986), что характерно для морей умеренных широт. Следовательно, широкая экологическая валентность и космополитизм этих видов способствует освоению новых акваторий, и их нахождение в перифитоне навигационных буев Морского заповедника вполне закономерно.
В перифитоне навигационных буев в бухтах Рейд Паллада, Экспедиции, Новгородская, Портовая, Порт-Посьет, Витязь отмечено более высокое видовое богатство (73 вида) диатомовых водорослей, многие из них имеют высокую температурную и соленосную толерантность, позволяющую им обитать в широком диапазоне факторов среды. В осенний период в большинстве мелководных бухт зал. Посьета отмечены максимальные концентрации нитритов, нитратов, фосфатов и кремния, являющихся важнейшими биогенными элементами, лимитирующими развитие диатомовых водорослей (Григорьева, 2012). Исследования различных групп макробентоса также свидетельствуют о том, что многие акватории зал. Посьета характеризуются преобладанием негативных индикаторов загрязнения, высоким видовым разнообразием бентофауны и являются на сегодняшний день одними из эталонов фоновых акваторий (Belan et al., 2003).
Важнейшим показателем экологического состояния акваторий зал. Посьета, прилегающих к Морскому заповеднику, являются виды-индикаторы органического загрязнения вод. Среди общего списка найденных видов-индикаторов 47% занимают β-мезосапробионты – индикаторы умеренного органического загрязнения вод и индикаторы «условно чистых» вод – ο-сапробионты (14%), ο-β-мезосапробионты (3%) и χ-ο-сапробионты (3%) (рис. 3). В импактной бух. Троицы видовое богатство (24 вида) диатомовых водорослей было почти в 3 раза ниже, чем в фоновых акваториях, а по числу видов в этой бухте преобладают индикаторы значительного органического загрязнения вод α-мезосапробионты (33%) над β-мезосапробионтами, что в значительной степени обусловлено хроническим воздействием антропогенного фактора со стороны морского порта Зарубино и рыбокомбината, находящегося на его территории. Среди α-мезосапробионтов виды диатомей H. cymbifera, T. fasciculata и M. moniliformis, по нашим данным, доминируют в наиболее загрязненных органикой бухтах зал. Петра Великого Японского моря (Бегун и др., 2010; Бегун, 2012).
Такая закономерность в значительной степени обусловлена тем, что неочищенные сточные воды, поступающие в бухту Троицы, а также нефтепродукты, попадающие в акваторию из морского порта, богатые биогенными соединениями, определяют развитие толерантных таксонов микроводорослей. Кроме того, в этой бухте, помимо нефтепродуктов, эпизодически происходит выброс в воду различных токсикантов, в том числе тяжелых металлов, значительно превышающих минимально допустимую концентрацию (Кобзарь, Христофорова, 2012) и оказывающих ингибирующее влияние на развитие большинства видов микроводорослей. В результате этого экологическая структура сообщества водорослей значительно упрощается, «аборигенные» виды замещаются «антропогенными», обладающими высокой толерантностью к загрязнению среды.
Кроме того, соседство крупного морского порта, в акватории которого происходит постоянный слив огромного объема неочищенного балласта с судов дальнего плавания (Белан, 2001), может оказать негативное влияние на биоту Морского заповедника. В результате деятельности морского транспорта (балластные воды и обрастание судов) следует также учитывать возможность переноса морских организмов и тот факт, что в зал. Петра Великого Японского моря действуют 2 вектора переноса видов – приазиатский и транстихоокеанский, представляющие собой потенциальную возможность переноса экзотических видов в акватории Морского заповедника (Звягинцев и др., 2012).
Заключение
В акваториях зал. Посьета Японского моря, прилегающих к Морскому заповеднику, исследован видовой состав и экология диатомовых водорослей перифитона на буях плавучего навигационного ограждения. В большинстве исследованных бухт (Рейд Паллада, Экспедиции, Новгородская, Портовая, Порт-Посьет, Витязь) в перифитоне отмечено высокое видовое богатство и преобладание видов β-мезосапробионтов, указывающих на умеренный уровень органического загрязнения вод. Для импактной бухты Троицы выявлено низкое видовое богатство перифитона и преобладание α-мезосапробионтов, свидетельствующие о высоком уровне органического загрязнения вод со стороны порта «Зарубино» и рыбокомбината, находящегося на его территории.
Говоря о перспективах успешного решения современных задач мониторинга биоты в морском заповеднике и прилегающих к нему акваторий, на начальном этапе необходимо создать научный фундамент – по возможности полную базу данных по видовому составу и количественному распределению его населения. Учитывая слабый уровень изученности микрофитобентоса этих акваторий, необходимо дальнейшее расширение флористических и гидробиологических исследований, включая мониторинг не только видового состава, но и данные по изучению сезонной динамики количественных показателей микроводорослей и анализу параметров водной среды.
Работа проведена при финансовой поддержке грантов РФФИ № 15-04-05643 А и № 14-34-50597 мол_нр.
Список литературы
1. Бегун А.А. Биоиндикация состояния морской среды по диатомовым водорослям эпифитона макрофитов (залив Петра Великого, Японское море) // Известия ТИНРО. 2012. Т. 169. С. 1-17.
2. Белан Т.А. Особенности обилия и видового состава бентоса в условиях загрязнения: Залив Петра Великого, Японское море: Автореф. дисс… канд. биол. наук. – Владивосток, 2001. – 24 с.
3. Герасимюк В.П., Эннан А.А., Шихалеева Г.Н. Видовой состав водорослей бентоса Куяльницкого лимана (северо-западное Причерноморье, Украина) // Альгология. 2011. 21, № 2. С. 226-240.
4. Григорьева Н.И. Залив Посьета: физико-географическая характеристика, климат, гидрологический режим / Современное экологическое состояние залива Петра Великого Японского моря: монография. Отв. ред. Н.К. Христофорова. – Владивосток: Издательский дом Дальневост. федерал. ун-та, 2012. – С. 31-61.
5. Гусляков Н.Е., Закордонец О.А., Герасимюк В.П. Атлас диатомовых водорослей бентоса северо-западной части Чёрного моря и прилегающих водоемов. – К.: Наук. думка, 1992. 112 с.
6. Дальневосточный морской биосферный заповедник. Биота. Т. II. /Отв. ред. А.Н. Тюрин, ред. А.Л. Дроздов. – Владивосток: Дальнаука, 2004. – 2. – 848 с.
7. Диатомовые водоросли СССР. Ископаемые и современные. Т. I. – Л.: Наука, 1974. – 400 с.
8. Звягинцев А.Ю., Ивин В.В., Кашин И.А., Бегун А.А., Городков А.Н. Чужеродные виды в Дальневосточном морском государственном биосферном заповеднике // Изв. ТИНРО. 2012. 170. С. 60–81.
9. Кобзарь А.Д., Христофорова Н.К. Оценка загрязнения вод залива Петра Великого тяжелыми металлами по их содержанию в бурых водорослях-макрофитах / Современное экологическое состояние залива Петра Великого Японского моря: монография. Отв. ред. Н.К. Христофорова. – Владивосток: Издательский дом Дальневост. федерал. ун-та, 2012. – С. 362–381.
10. Лоция северо-западного берега Японского моря. От реки Туманная до мыса Белкина. – Министерство обороны СССР. Гл. управление навигации и океанографии. – 1984. – 320 с.
11. Малютин А.Н., Гульбина А.В. Дальневосточных морской биосферный заповедник ДВО РАН: история создания, биота, современное состояние / Современное экологическое состояние залива Петра Великого Японского моря: монография. Отв. ред. Н.К. Христофорова. – Владивосток: Издательский дом Дальневост. федерал. ун-та, 2012. – С. 175-190.
12. Николаев В.А. Вертикальное распределение бентосных диатомовых водорослей в заливе Посьета // Прибрежные сообщества дальневосточных морей. 1976. № 6. С. 94–98.
13. Оксиюк О.П., Давыдов О.А. Методологические принципы оценки состояния водных объектов по микрофитобентосу // Гидробиол. журн. 2006. 42, № 2. С. 98-112.
14. Рябушко Л.И. Диатомовые водоросли верхней сублиторали северо-западной части Японского моря // Автореф. дисс. ... канд. биол. наук: Севастополь. – 1986. – 24 с.
15. Рябушко Л.И. Микрофитобентос Черного моря. – Севастополь: ЭКОСИ–Гидрофизика, 2013. – 416 с.
16. Belan T.A., Tkalin A.V., Lishavskaya T.S. The present status of bottom ecosystems of Peter the Great Bay (the Sea of Japan) // Pacific Oceanography. 2003. 1, № 2. P. 158–167.
17. Truby E.W. Preparation of single-celled marine dinoflagellates for electron microscopy // Microscopy Research and Technique. 1997. Vol. 36. P. 377-340.
опубликовано - декабрь 2014 г.
Ecology of periphyton diatoms of specially protected natural territories for example to Far-Eastern Marine State Natural Biosphere Reserve (Posyet Bay, Japan Sea)
Andrey A. Begun
Species composition and ecology characteristic of periphyton diatoms on navigation buoys in the water area of Posyet Bay, Japan Sea are investigated. It is shown that for some water areas of Posyet Bay characterized by the dominance of significant organic pollution indicators under background low levels of number species in diatom periphyton. Features of species composition and ecology of periphyton diatoms is largely determined by the level of anthropogenic pollution of water areas, adjacent to Marine reserve.
Key words: periphyton; diatoms; sapropity; navigation buoys; Marine reserve; Posyet Bay; Japan Sea.
Об авторе
Бегун Андрей Аркадьевич - Begun Andrey A.
кандидат биологических наук
научный сотрудник, Лаборатория экологии шельфовых сообществ, Институт биологии моря им.А.В.Жирмунского ДВО РАН, г. Владивосток, Россия (Zhirmunsky Institute of Marine Biology, Far Eastern Branch, Russian Academy of Sciences, Vladivostok, Russia)
andrejbegun@yandex.ru
Корреспондентский адрес: Россия, 690041, г. Владивосток, ул. Пальчевского, д. 17; телефон (423) 231-09-12.
ССЫЛКА НА СТАТЬЮ:
Бегун А.А. Экология диатомовых водорослей перифитона особо охраняемых природных территорий на примере Дальневосточного морского государственного природного биосферного заповедника (залив Посьета, Японское море) // Вопросы современной альгологии. 2014. № 2 (6). URL: http://algology.ru/558
Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно.
Адрес - info@algology.ru
При перепечатке ссылка на сайт обязательна
К другим статьям Международной конференции «Экологическая физиология водных фототрофов: распространение, запасы, химический состав и использование». III Сабининские чтения.
.
Карта сайта

|
