из Материалов Международной конференции «Экологическая физиология водных фототрофов: распространение, запасы, химический состав и использование» III Сабининские чтения 29 ноября - 29 декабря 2014 г.
Водоросли во льду и в воде Белого моря
в переходный период от зимы к весне
Algae in the ice and water of the White Sea
in winter–spring transition period
Сажин А.Ф.1, Житина Л.С.2, Сергеева В.М.1,
Ратькова Т.Н.1, Бахмет И.Н.3
Andrey F. Sazhin, Ludmila S. Zhitina, Valentina М. Sergeeva,
Tatiana N. Rat’kova, Igor N. Bakhmet
1Институт океанологии им. П.П.Ширшова РАН
2Московский государственный университет им. М.В.Ломоносова,
биологический факультет
3Институт биологии Карельского научного центра РАН
УДК 574.52 + 581.526.325.132
Описана структура планктонного фитоцена прибрежной зоны Белого моря в переходный период от зимы к весне. Как в подледном фитопланктоне во время интенсивного таяния льда, так и в воде в последующий период весны и начала лета, среди автотрофных форм преобладают диатомовые водоросли, не являющиеся массовыми во льду. Миксотрофные и гетеротрофные динофлагелляты могут преобладать среди водорослей вплоть до конца июня.
Ключевые слова: ледовые и планктонные водоросли; Белое море; весна; автотрофные, миксотрофные и гетеротрофные формы.
Введение
В водных морских экосистемах высоких широт активность организмов всех уровней, в том числе фитопланктона и ледовых водорослей, наиболее высока в весенние, летние и осенние месяцы, а наиболее низкая – в зимний период. Что же касается наиболее яркого переходного периода от зимы к весне с резкими изменениями температурного, светового, гидродинамического, гидрохимического и других режимов, тут в наших знаниях существуют значительные пробелы. Особенно важно иметь представление о количественных характеристиках и активности водорослей в период вскрытия водоемов ото льда. В это время интенсивно функционируют микробные пищевые сети, и основные потоки энергии проходят не только через автотрофный, главным образом диатомовый, компонент, но и через гетеротрофные и миксотрофные водоросли, преимущественно перидинеи. Тем не менее, даже для наиболее доступного для исследований Белого моря в межсезонье мы имеем лишь немногочисленные данные, полностью представленные в обзорных работах (Ильяш и др., 2012 а, Ильяш и др., 2012 б; Сажин и др., 2012). Диатомеи – группа, дающая основной вклад в суммарную биомассу ледовых организмов. Иногда водоросли могут достигать такой высокой концентрации, что нижняя часть льда заметно окрашена в коричневый цвет. Численность клеток диатомей в таком «окрашенном» слое льда составляет более 100 тыс. кл./л. В состав доминирующих видов входят как типичные происходящие из планктона или бентоса ледовые формы (Nitzschia frigida, Entomoneis kjellmanii, E. paludosa, Navicula pelagica, Pauliella taeniata, Tryblionella gracilis и др.), так и более редкие планктонные (Thalassiosira gravida, T. nordenskioeldii и др.), а иногда и преимущественно бентосные (Amphora spp., Cocconeis spp. и др.). Набор доминирующих форм может меняться от района к району и от года к году, то есть имеет большую пространственно-временную вариабельность (Сажин и др., 2012). В Белом море практически вся биомасса, образовавшаяся в результате «цветения» ледовых диатомей, попадает в воду при таянии льда и может служить основой пелагических пищевых цепей задолго до начала интенсивного размножения весеннего фитопланктона. Ряд ледовых видов развивается в верхнем слое воды, формируя первую фазу «цветения» водорослей. Основная же масса попадающих в воду ледовых диатомей, по-видимому, довольно быстро потребляется планктонными гетеротрофами непосредственно подо льдом. В развивающемся после таяния льда «цветении» водорослей, как правило, доминируют типичные планктонные виды, не являющиеся массовыми во льду. Исследованию фитопланктона именно в периоды зимне-весеннего межсезонья, когда биота формируется в новых окружающих условиях, происходящих со сменой всей структуры среды обитания, и посвящена данная работа.
Материал и методика
Для решения поставленных задач в одной и той же точке губы Чупа Кандалакшского залива Белого моря (в районе Беломорской биологической станции «Картеш» ЗИН РАН) были отобраны пробы фитопланктона (Ст. 5, рис. 1).
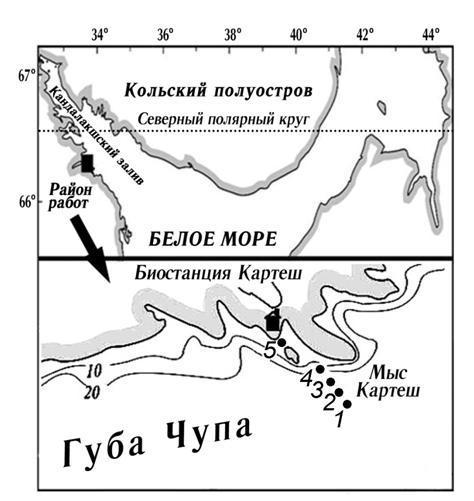
Рис. 1. Карта-схема района работ
Материал был собран вручную из поверхностного слоя моря при максимальном приливе 3,7,11,15 и 19 апреля 2011 г., а также с 11 марта по 24 июня 2012 г. (25 отборов с интервалом 3-5 дней). Объем пробы составлял 200-250 мл, пробы фиксировали раствором Люголя с добавлением глютарового диальдегида. Численность водорослей определяли методом прямого счета под световым микроскопом в камерах Ножотта (объем 0,1 мл) и Наумана (объем 1 мл). Трофический статус водорослей определяли на основании собственных многолетних данных люминесцентной микроскопии, полученных в районе исследований, и по литературным данным. Специально отметим, что в отличие от подавляющего большинства автотрофных диатомей, практически все динофлагелляты являются миксотрофами или гетеротрофами (Stoecker, 1999; Jeong et al., 2010). Миксотрофные динофлагелляты способны и к фотосинтезу и к фаготорофии. Добычей могут служить бактерии и разнообразные простейшие, включая водоросли, а также яйца и младшие стадии многоклеточных животных (Jacobson, Anderson 1996; Stoecker 1999; Jeong et al. 2005, 2008, 2010; Seong et al. 2006).
Величину биомассы в углеродных единицах рассчитывали исходя из объемов клеток (Menden-Deuer, Lessard, 2000).
В работе мы также использовали собственные данные по ледовым водорослям, собранные в этом же районе на станциях 1-4 в 2002-2012 гг. (рис. 1). Ледовые керны были отобраны с помощью титанового бура и разделены для анализов на 3 или более частей в зависимости от структуры льда. Подробная методика планктонных и ледовых работ, а также подготовки проб к микроскопии изложена в работе Сажина с соавт. (2012).
Результаты
В конце марта - начале апреля 2011 г. дневная температура воздуха в районе ББС ЗИН «Картеш» составляла -2-6° С, а ночная опускалась до -7-11° С. Температура воды в подледном слое в это время не превышала -1,2-1,3° С. К 20 апреля днем воздух прогревался до +5-7° С, а температура подледного слоя воды колебались вокруг нулевого значения. Акватория губы Чупа в районе исследований в конце апреля 2011 г. была покрыта льдом.
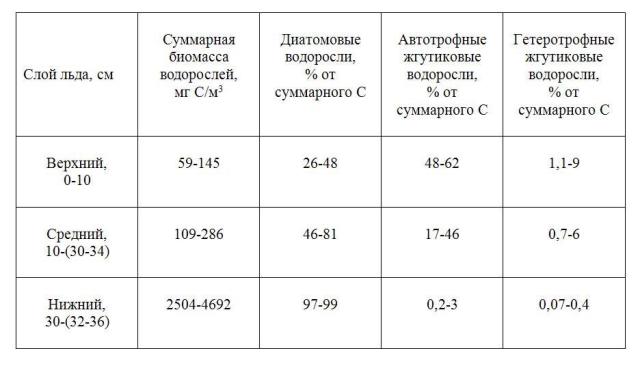
Судя по нашим данным для апреля как 2002, так и 2011 гг. во время интенсивного таяния льда основная масса ледовых водорослей сосредоточена в его нижнем слое и представлена преимущественно диатомеями, среди которых доминируют Nitzschia frigida, Entomoneis kjellmanii и E. paludosa. Иногда существенный вклад в суммарную численность и биомассу ледовой флоры могут вносить Tryblionella gracilis и Thalassiosira nordenskioeldii. Вклад автотрофных диатомей в суммарную биомассу водорослей составляет 97-99%. Доля автотрофных и гетеротрофных жгутиковых водорослей в нижних слоях льда не превышает 3% суммарного углерода ледовой альгофлоры (Таблица 1). Миксотрофные формы практически отсутствуют.
В подледном фитопланктоне в апреле 2011 г. преобладали представители рода Navicula. Общая численность Navicula spp. в воде в период 3-19 апреля 2011 г. варьировала в пределах 930-21560 кл./л, биомасса составляла 0,19-0,75 мгС/м3. Самым массовым видом среди навикул была N. pelagica. Концентрация Nitzschia frigida не превышала 200-980 кл./л, или 0,004-0,03 мгС/м3. Заметный вклад в суммарную численность и биомассу диатомей вносили лишь Pauliella taeniata, Chaetoceros tenuissimus, Cocconeis scutellum. Численность всего подледного диатомового компонента менялась от 8300 до 110500 кл./л, биомасса составляла 0,55-3,61 мгС/м3 (рис. 2 а). Практически такие же величины относятся и ко всем планктонным автотрофным водорослям. Явных признаков «цветения» диатомей в апреле 2011 г. мы не наблюдали.
Таблица 1. Вклад различных групп водорослей (%) в их суммарную биомассу (мгС/м3) в разных слоях льда Белого моря во время интенсивного таяния ледового покрова.
Среди миксотрофного и гетеротрофного фитопланктона преобладали молодые Amphidiniumm spp., Eutreptiella braarudii, Gyrodinium spirale. С 3 по 19 апреля общая численность этих водорослей составляла не более 2500-3000 кл./л, а суммарная биомасса не превышала 1 мгС/м3. В этот период самыми многочисленными были споры перидиней, концентрация которых доходила до 10 тысяч кл./л, при биомассе до 1,5 мгС/м3. Суммарная численность миксотрофного и гетеротрофного фитопланктона варьировала в пределах 2450-14000 кл./л, а биомасса составляла 1,1-2,3 мгС/м3 (рис. 2 б).
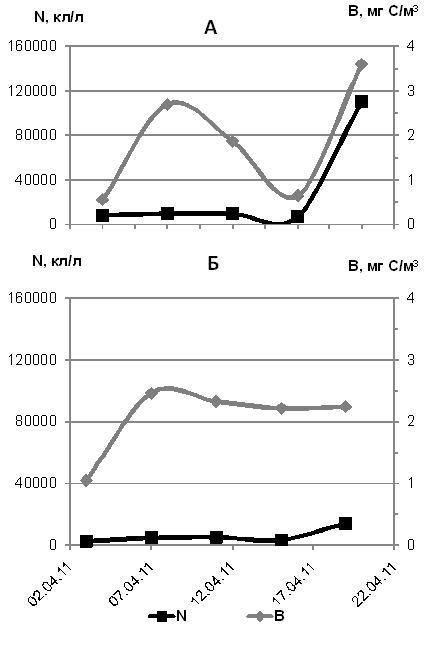
Рис. 2. Динамика численности (N, кл/л) и биомассы (B, мг С/м3) автотрофных (диатомовых) водорослей (А), миксотрофных и гетеротрофных водорослей (Б) в апреле 2011 г.
Доля автотрофных представителей альгофлоры, преимущественно диатомей, в период исследований 2011 г. варьировала в пределах 66-89% (в среднем 73%) суммарной численности автотрофных, миксотрофных и гетеротрофных водорослей, а их биомасса составляла 23-62% (в среднем 43%) суммарной биомассы автотрофов, миксотрофов и гетеротрофов. Миксотрофные и гетеротрофные водоросли в этот период в среднем составляли, соответственно, 27% суммарной численности и 57% биомассы всего фитопланктона (рис. 3 а,б).
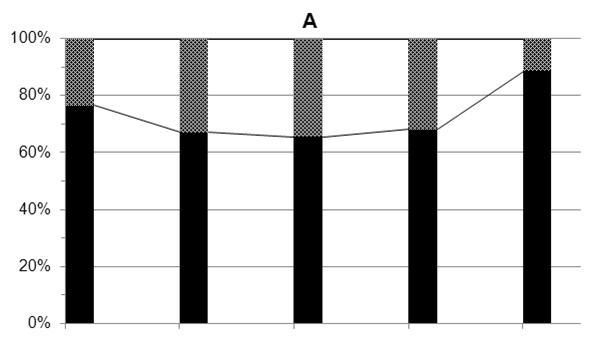
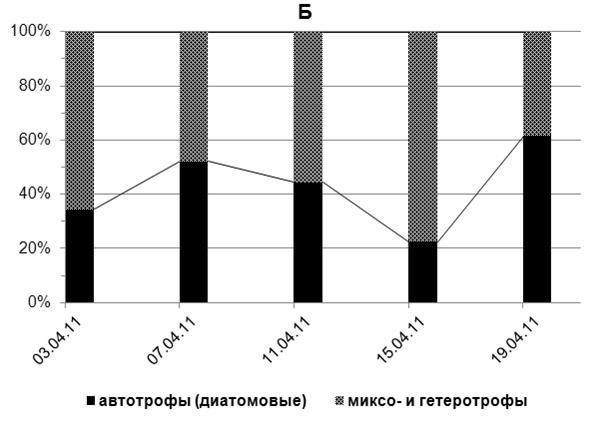
Рис. 3. Доля (%) автотрофных (диатомовых), миксо- и гетеротрофных водорослей в суммарной численности (А) и суммарной биомассе (Б) фитопланктона в апреле 2011 г.
В 2012 г. весь март и апрель температура воды в подледном слое в районе ББС ЗИН «Картеш» не превышала -1,4° С, несмотря на положительные значения температуры воздуха днем, а то и в ночные часы. Район исследований полностью освободился ото льда лишь к середине мая.
Судя по данным 2012 г., представители рода Navicula, как и в 2011 г., доминировали среди диатомей в течение всего исследуемого периода, уступая первенство Nitzschia frigida лишь в начале мая. Суммарная численность Navicula spp. в марте не превышала 1325 кл./л (0,24 мгС/м3), в апреле 5120 кл./л (1,28 мгС/м3), 3 мая обилие этих водорослей было максимальным (35970 кл./л или 3,82 мгС/м3), но к 7-11 мая составляло всего 4580-4710 кл./л (0,39-0,51 мгС/м3) и далее, вплоть до конца июня, не превышало 616 кл./л или 0,09 мгС/м3. Nitzschia frigida встречалась в планктоне в небольшом количестве вплоть до конца апреля и лишь в начале мая ее численность возросла до величины 79570 кл./л (5,91 мгС/м3), что привело к доминированию этого вида среди диатомей. Уже к концу первой декады мая концентрация N. frigida снизилась на порядок, и после 8 июня этот вид полностью отсутствовал в наших пробах. Entomoneis spp. в небольшом количестве присутствовали в планктоне лишь в середине марта, конце апреля и начале мая. Pauliella taeniata встречалась в планктоне крайне редко, в виде отдельных клеток. Представители рода Chaetoceros присутствовали лишь в пробах от 28 апреля, 19 мая и 20-24 июня. Cocconeis scutellum впервые был отмечен 28 апреля и в более поздний период заметной роли не играл.
Численность всех диатомовых водорослей марте-июне 2012 г. менялась от 265 до 135000 кл./л, максимальное значение биомассы составляло 12 мгС/м3 (рис. 4а). Такими же величинами характеризовались численность и биомасса всех планктонных автотрофных водорослей. Наиболее интенсивное развитие диатомей мы наблюдали в начале мая, когда пик концентрации Navicula spp. сменился максимумом обилия Nitzschia frigida.
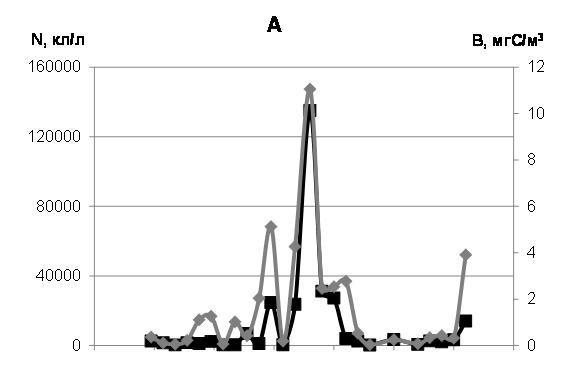
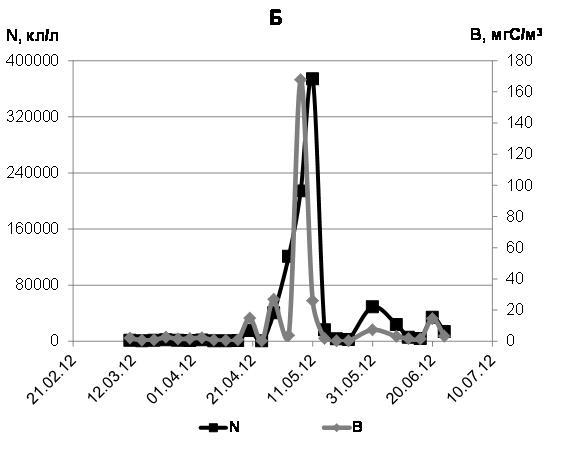
Рис. 4. Динамика численности (N, кл/л) и биомассы (B, мг С/м3) автотрофных (диатомовых) водорослей (А), миксотрофных и гетеротрофных водорослей (Б) в марте-июне 2012 г.
Среди миксотрофного и гетеротрофного фитопланктона в марте-июне 2012 г. преобладала Peridiniella catenata (c максимумом обилия до 170 тысяч кл./л при биомассе 166 мгС/м3 в начале мая), заметную роль играли Dinophysis arctica, Prorocentrum balticum и представители рода Protoperidinium.
В мае 2012 г. в планктоне были необычайно многочисленны миксотофные Euglena spp. при доминировании E. acusformis. 11 мая мы наблюдали максимум обилия этих водорослей. Их численность доходила до 300 тысяч кл./л при биомассе 17,32 мгС/м3. Споры же перидиней в период исследований 2012 г. заметной роли в планктоне не играли. Суммарная численность миксотрофного и гетеротрофного фитопланктона варьировала в пределах 400-374000 кл./л, а биомасса в десятых числах мая, в период максимального развития миксотрофов и гетеротрофов, доходила до 167 мгС/м3 (рис. 4 б).
Доля автотрофных водорослей, преимущественно диатомей, в период исследований 2012 г. варьировала от 2 до 92% (в среднем 38%) суммарной численности автотрофных, миксотрофных и гетеротрофных клеток, а их биомасса составляла 3-75% (в среднем 28%), соответственно, суммарной биомассы автотрофов, миксотрофов и гетеротрофов. Миксотрофные и гетеротрофные водоросли в этот период в среднем составляли, соответственно, 62% и 72%, суммарной численности и биомассы всех водорослей, то есть практически преобладали в фитопланктоне начиная с середины марта и вплоть до третьей декады июня 2012 г. (рис. 5 а, б).
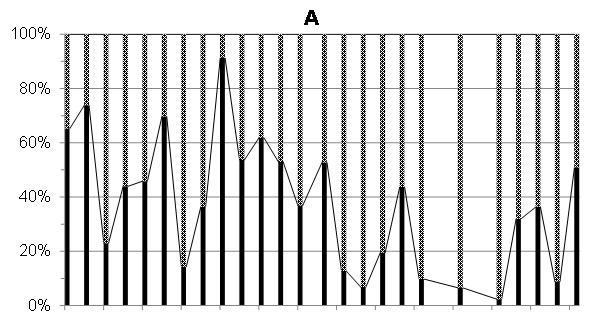
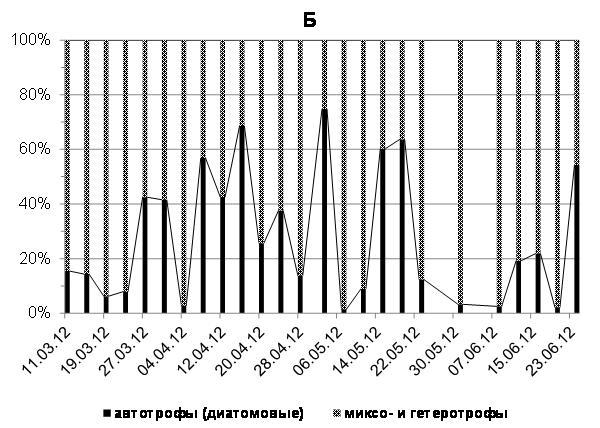
Рис. 5. Доля (%) автотрофных (диатомовых), миксо- и гетеротрофных водорослей в суммарной численности (А) и суммарной биомассе (Б) фитопланктона в марте-июне 2012 г.
Заключение
Собранный нами в 2011 и 2012 гг. материал не позволил в полной мере учесть самые мелкие формы водорослей. Тем не менее, судя по нашим данным, полученным в другие годы в сезон таяния льда в этом же месте, подледный фитоцен в эти периоды имеет сходную структуру. До 50% в суммарную биомассу водорослей в разных слоях подледной воды могут вносить гетеротрофные жгутиковые и не более 60% – автотрофные жгутиковые формы (Ратькова и др., 2004).
Таким образом, как в подледном фитопланктоне во время интенсивного таяния льда, так и в воде в последующий период весны и начала лета, среди автотрофных форм преобладают диатомовые водоросли, не являющиеся массовыми во льду. По мере наступления безледного периода происходит смена доминирующих форм, и в разные годы набор массовых видов диатомовых водорослей может сильно различаться. Лишь Nitzschia frigida – один из наиболее массовых видов ледовых диатомей, продолжает успешно существовать в планктоне, достигая значительного обилия вплоть до середины мая.
Как мы видели, миксотрофные и гетеротрофные формы могут преобладать среди водорослей вплоть до конца июня. В самом начале весны их жизнедеятельность обеспечивается поступающим из тающего льда в воду органическим веществом, главным образом, за счет диатомовых водорослей из нижних слоев ледового покрова. Миксотрофные и гетеротрофные динофлагелляты способны не только усваивать органические соединения, но и питаться бактериями, пико- и нанопланктоном, включая жгутиковые формы, диатомей, других динофлагеллят и простейших, а также многоклеточные организмы. Такая возможность обеспечивается наличием у динофлагеллят самых разнообразных механизмов добычи и потребления пищи (Jeong et al., 2010). То есть, особенности жизнедеятельности миксотрофных и гетеротрофных динофлагеллят еще более усложняют существующие схемы планктонных пищевых сетей и заставляют нас по-новому оценивать роли этих водорослей в функционировании пелагических экосистем.
Работа выполнена при поддержке грантов РФФИ № 12-04-93081-Норв(а) и 14-05-00028.
Список литературы
1. Ильяш Л.В., Ратькова Т.Н., Радченко И.Г., Житина Л.С., Белевич Т.А., Федоров В.Д. Состав и пространственно-временная изменчивость фитопланктона // Биологические ресурсы Белого моря: изучение и использование. Исследования фауны морей. Ред.: Бергер В.Я. - Т.69 (77). - СПб: ЗИН РАН, 2012а. - С. 48-67.
2. Ильяш Л.В., Житина Л.С., Кудрявцева В.А., Мельников В.А. (2012б) Сезонная динамика видового состава и биомассы водорослей в прибрежных льдах Кандалакшского залива Белого моря // Журнал общей биологии. 2012б. Том 73. № 6. С. 459-470.
3. Ратькова Т.Н., Сажин А.Ф., Кособокова К.Н. Одноклеточное население беломорской пелагиали подо льдом в ранне-весенний период // Океанология. 2004. Том 44. № 2. С. 259-266.
4. Сажин А.Ф., Ратькова Т.Н., Мошаров С.А, Романова Н.Д., Мошарова И.В., Портнова Д.А. Биологические компоненты сезонного льда // Биологические ресурсы Белого моря: изучение и использование. Исследования фауны морей. Ред.: Бергер В.Я. - Т.69 (77). - СПб: ЗИН РАН, 2012. - С. 97-115.
5. Menden-Deuer S., Lessard E.J. Carbon to volume relationship for dinoflagellates, diatoms and other protists plankton // Limnol. Oceanogr. 2000. V. 45. № 3. P. 569-579.
6. Jacobson D.M., Anderson D.M. Widespread phagocytosis of ciliates and other protists by marine mixotrophic and heterotrophic thecate dinoflagellates // J Phycol. 1996. V. 32. P. 279-285.
7. Jeong H.J., Yoo Y.D., Park J.Y., Song J.Y., Kim S.T., Lee S.H., Kim K.Y.,Yih W.H. Feeding by the phototrophic red-tide dinoflagellates: five species newly revealed and six species previously known to be mixotrophic // Aquat. Microb Ecol. 2005. V. 40. P.133-155.
8. Jeong H.J., Seong K.A., Yoo Y.D., Kim T.H., Kang N.S., Kim S., Park J.Y., Kim J.S., Kim G.H., Song J.Y. Feeding and grazing impact by small marine heterotrophic dinoflagellates on hetertrophic bacteria // J. Eukaryot. Microbiol. 2008. V. 55. P. 271-288.
9. Jeong H.J., Yoo Y.D., Kim J.S., Seong K.A., Nam Kang N.S., Kim T.H. Growth, Feeding and Ecological Roles of the Mixotrophic and Heterotrophic Dinoflagellates in Marine Planktonic Food Webs // Ocean Sci. J. 2010. V. 45. № 2. P. 65-91.
10. Seong K.A., Jeong H.J., Kim S., Kim G.H., Kang J.H. Bacterivory by co-occurring red-tide algae, heterotrophic nanoflagellates, and ciliates on marine bacteria // Mar. Ecol. Prog. Ser. 2006. V. 322. P. 85-97.
11. Stoecker D.K. Mixotrophy among Dinoflagellates // J. Eukaryot. Microbiol. 1999. V. 46. № 4. P. 397-401.
опубликовано - ноябрь 2014 г.
Algae in the ice and water of the White Sea in winter–spring transition period
Andrey F. Sazhin, Ludmila S. Zhitina, Valentina М. Sergeeva, Tatiana N. Rat’kova, Igor N. Bakhmet
The highest life activity of all organisms, including plankton Algae, in the polar coastal ecosystems, comes during spring, summer and autumn, and the lowest one – in winter period. But we have poor knowledge about structure and functioning of water ecosystems during the brightest winter-spring transition period. This period («offseason») is characterized by sharp variations of temperature, illuminance, hydrodinamic and hydrochemical conditions. It makes very important to have an idea and understanding about quantity and activity of Algae during the ice melting period and just after it. Microbial food web and microbial «loop» are very active in this time, and main energy flow comes also through mixotrophic and heterotrophic Dinoflagellates. This investigation deals with Algae from ice and plankton in winter-spring transition period, when biota is forming under new surrounding conditions. All structure of biota is changing in this period.
Key words: ice and plankton Algae; White Sea; winter-spring transition period; phototrophic; mixotrophic and heterotrophic species.
Об авторах
Сажин Андрей Федорович – Sazhin Andrey Fiodorovich
кандидат биологических наук
старший научный сотрудник Института океанологии им. П.П. Ширшова РАН, Москва, Россия (P.P.Shirshov Institute of Oceanology RAS, Moscow, Russia)
andreysazhin@yandex.ru
Житина Людмила Сергеевна – Zhitina Ludmila Sergeevna
кандидат биологических наук
старший научный сотрудник ФГБОУ ВПО «Московский государственный университет им.М.В.Ломоносова», Москва, Россия (Lomonosov Moscow State University, Moscow, Russia), Биологический факультет, кафедра гидробиологии
lgitina@mail.ru
Сергеева Валентина Михайловна – Sergeeva Valentina Mikhailovna
кандидат биологических наук
научный сотрудник Института океанологии им. П.П. Ширшова РАН, Москва, Россия (P.P.Shirshov Institute of Oceanology RAS, Moscow, Russia)
vsergeeva@gmail.com
Ратькова Татьяна Николаевна – Rat’kova Tatiana Nikolaevna
кандидат биологических наук
старший научный сотрудник Института океанологии им. П.П. Ширшова РАН, Москва, Россия (P.P.Shirshov Institute of Oceanology RAS, Moscow, Russia)
tatiana.ratkova@mail.ru
Бахмет Игорь Николаевич – Bakhmet Igor Nikolaevich
кандидат биологических наук
старший научный сотрудник Института биологии Карельского Научного Центра РАН, Петрозаводск, Россия (Institute of Biology Karelian Research Centre RAS, Petrozavodsk, Russia)
igor.bakhmet@gmail.com
Корреспондентский адрес: Россия, Москва, 117997, Институт океанологии РАН, Нахимовский проспект, д. 36; тел. 8(499)124-85-26.
ССЫЛКА НА СТАТЬЮ:
Сажин А.Ф., Житина Л.С., Сергеева В.М., Ратькова Т.Н., Бахмет И.Н. Водоросли во льду и в воде Белого моря в переходный период от зимы к весне // Вопросы современной альгологии. 2014. № 2 (6). URL: http://algology.ru/522
При перепечатке ссылка на сайт обязательна
К другим статьям Международной конференции «Экологическая физиология водных фототрофов: распространение, запасы, химический состав и использование». III Сабининские чтения.
.
Карта сайта

|
