
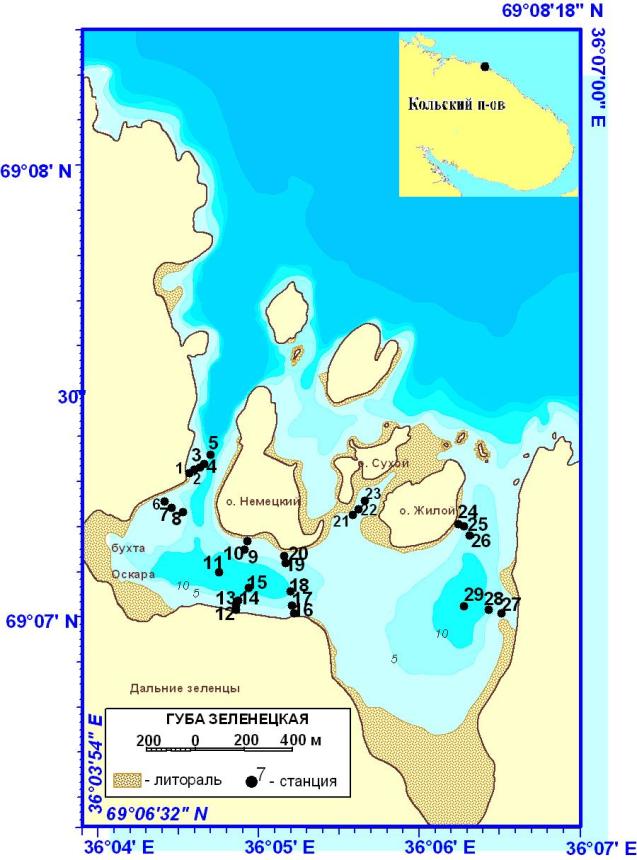
|
|
|
Структура сублиторальных фитоценозов защищенной губы Зеленецкая Мурманского побережья Баренцева моря
|
|||||
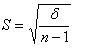 |
, где δ – дисперсия выборки, n-1 – число степеней свободы. |
Оценивали встречаемость каждого вида как долю в процентах непустых проб (Василевич, 1969). Выравненность видов по обилию на каждой станции оценивали по средней доле доминантного и субдоминантного видов, а также индексам видового разнообразия Шеннона и Симпсона (Лебедева и др., 2004). Проверку на нормальность распределения данных проводили через критерий χ-квадрат. Также был выполнен корреляционный анализ зависимости видового разнообразия (значений индекса Шеннона) от глубины, грунта и вида-доминанта.
Фитоценозы дифференцировали по принципу однородности видового состава, пространственной структуры и приуроченности к определенным условиям. Сходство видового состава определяли с применением меры сходства Брея-Куртиса после логарифмического преобразования данных по биомассе различных видов методом попарного присоединения (Field et al., 1982).
Расчеты выполняли в программах Excel 2003 и PAST (Hammer et al., 2008).
Результаты и обсуждение
Видовой состав макрофитобентоса и распределение водорослей
В сублиторальной зоне губы Зеленецкая было обнаружено 67 видов водорослей-макрофитов, из них 23 вида бурых водорослей, 29 красных и 15 зеленых (табл. 1 a, b, c). Только 7 видов водорослей распределяются по губе относительно равномерно (были обнаружены более чем в 60% проб). Многие водоросли с биомассой превышающей 0,1 г/м², такие как Saccorhyza dermatodea, Halosiphon tomentosus, Ulva lactuca L. и другие (табл. 1 a, b, c), отмечены в пробах единично, поэтому выявить закономерности их распределения или приуроченности к определенным условиям обитания не представляется возможным.
Таблица 1a. Биомасса и встречаемость макрофитов в губе Зеленецкая (бурые водоросли)
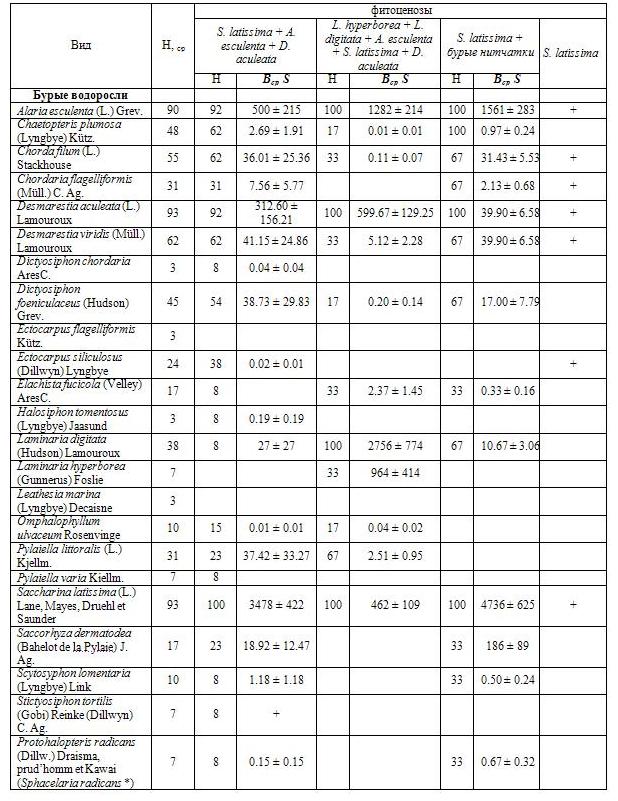
Таблица 1b. Биомасса и встречаемость макрофитов в губе Зеленецкая (красные водоросли)

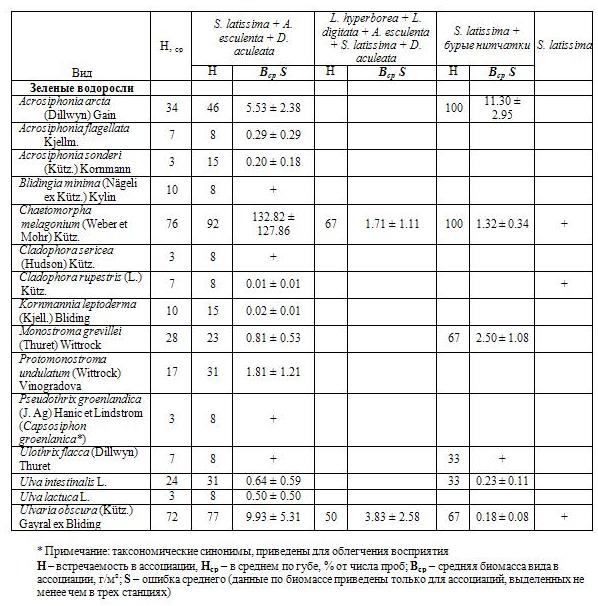
Таблица 1c. Биомасса и встречаемость макрофитов в губе Зеленецкая (зеленые водоросли)
Распределение водорослей по губе неравномерное. В зависимости от типа грунта проективное покрытие дна водорослями может составлять от 0 до 100% (табл. 2). Средняя биомасса свыше 1 г/м² характерна для 23 видов, еще у 17 видов она составляла более 0,1 г/м².
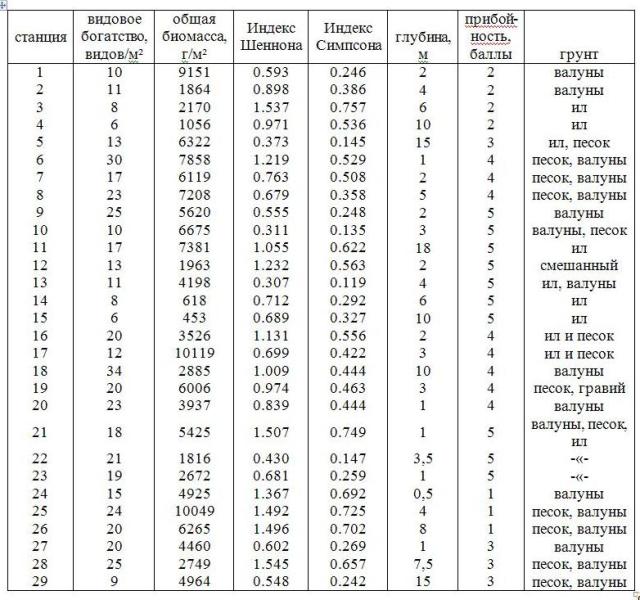
Сублиторальным фитоценозам губы свойственны низкие показатели выравненности видов по обилию (выраженному биомассой вида на станции), которая описывается логнормальным распределением, что характерно для растительных сообществ (Суханов, Жуков, 2003). Основная биомасса приходится на бурые водоросли. Доля доминантов (S. latissima, L. digitata, A. esculenta и D. aculeata) на порядок выше всех остальных. Видовое разнообразие варьирует в широких пределах: значения индекса Шеннона в исследованных сообществах колебались от 0,307 до 1,545, индекса Симпсона – от 0,119 до 0,757 (табл. 2). Зависимости видового разнообразия от глубины, типа грунта или доминирующего вида не выявлено.
Таблица 2. Количественные характеристики станций
По вертикали сублиторальные фитоценозы защищенной части губы Зеленецкая разделяются на три яруса: верхний ярус крупных многолетних ламинариевых, средний ярус одно-двухлетних макрофитов меньшего размера и нижний ярус корковых известковых водорослей (эпизодично). Наиболее явно ярусы выделяются в фитоценозах, подверженных воздействию прибоя. Синузии эпифитов обнаружены на стволиках и ризоидах ламинариевых водорослей, а также на Desmarestia aculeata и Chaetomorpha melagonium.
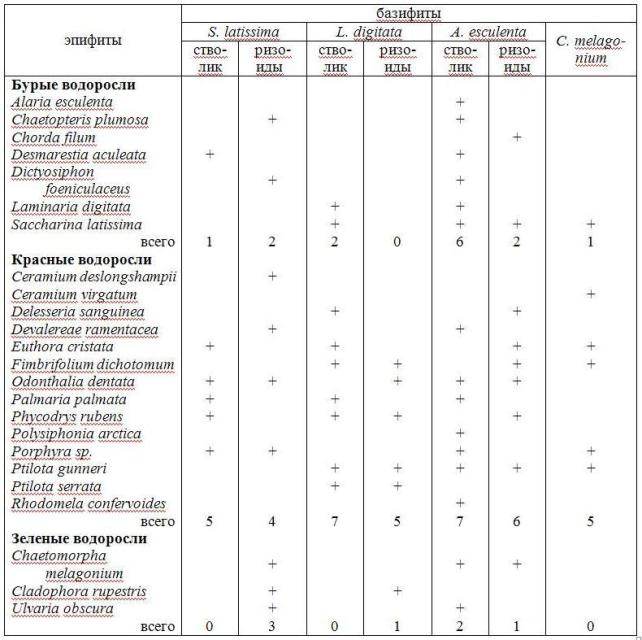
Видовой состав синузий эпифитов многолетних видов (S. latissima, A. esculenta и L. digitata) очень сходен, преобладают красные водоросли (табл. 3). Частично эти же виды были обнаружены и на ряде талломов C. melagonium, на которой также часто встречается Euthora cristata (C. Ag.) J. Ag., отмечены Ceramium virgatum Roth., Fimbrifolium dichotomum (Lepechin) Hansen, Ptilota gunneri Silva, Maggs, Irvine и проростки Porphyra sp. и Laminaria sp. Причем масса эпифитов, произрастающих на C. melagonium, часто превышала ее собственную. На D. aculeata единично отмечены проростки ламинариевых (трудно определяемы до вида, но предположительно S. latissima), Porphyra sp. (молодые стерильные особи). Все указанные нами виды эпифитов произрастают также и на грунте. Обнаружено только два вида облигатных эпифитов ламинариевых: Peyssonnelia sp. Decaisne и Rhodophysema elegans (Crouan, Crouan ex J. Ag.) Dixon.
Таблица 3. Видовой состав синузий эпифитов в защищенной части губы Зеленецкая
В результате кластеризации станций на уровне сходства свыше 40% было выделено 5 групп, в которые вошли 24 станции. На остальных 5 станциях видовой состав индивидуален (рис. 2).
1. Наиболее крупная группа (13 станций) выделяется на 50% уровне сходства. Этот кластер отличается большим числом видов, высокой долей и разнообразием зеленых водорослей. Разнообразие бурых водорослей сравнительно невелико.
2. Вторая группа состоит из 4 станций (21, 24-26) с уровнем сходства 54%, расположенных в проливах между островами. Во всех пробах отмечены красные водоросли Euthora cristata, Odonthalia dentata, Ptilota gunneri, Fimbrifolium dichotomum, Phycodrys rubens, Polysiphonia arctica и зеленые водоросли Chaetomorpha melagonium и Ulvaria obscura.
3. Третья группа из трех станций (17, 19, 20), расположенных между о. Немецкий и берегом, объединяется на уровне сходства 47%.
4. Кластер из станций 14 и 15, расположенных на глубинах 6-10 м в защищенной части губы, отличается бедностью видового состава. Степень сходства очень высокая, почти 80%.
5. Второй «бедный» видами кластер составляют станции 3 и 4, расположенные в западном проливе, также на илистых грунтах на глубине 6 и 10 м. Степень сходства только 47%.
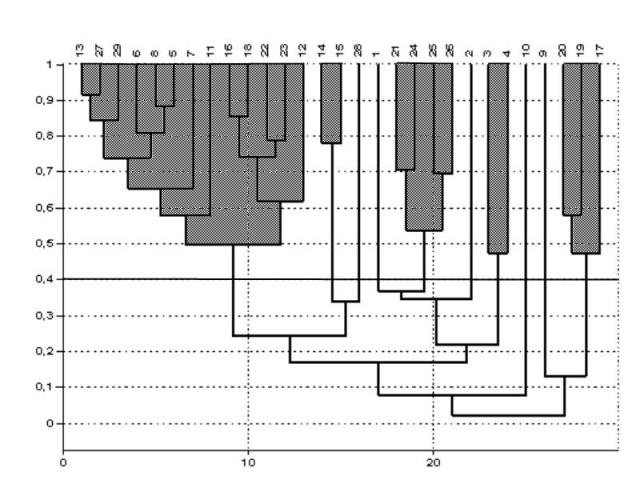
Рис. 2. Дендрограмма сходства фитоценозов губы Зеленецкая. Числа вверху – номера станций.
Пять станций не вошли ни в одну группу. Фитоценоз на ст. 9 сходен с первой группой по доминанту, но значительно отличается по составу видов. Станции 1 и 2 близки по составу массовых видов к фитоценозам второй группы, однако отличаются от них меньшей биомассой и разнообразием видов второго яруса. На станции 10 доминирует D. aculeata, как и на ст. 4, но биомасса водорослей значительно выше, заросли сомкнутые. Станция 28 отличается от остальных по видовому составу. Здесь доминирует S. dermatodea, обильны Porphyra ssp., U. obscura, Desmarestia viridis, Alaria esculenta. Для выделения вышеперечисленных станций в отдельные фитоценозы данных недостаточно.
В результате анализа полученных данных в губе Зеленецкая было выделено 4 фитоценоза (рис. 3):
1. Фитоценоз S. latissima + A. esculenta + D. aculeata с невыраженной ярусностью и средней биомассой синузий эпифитов и видов второго яруса, однако с высоким видовым разнообразием. Расположен в защищенных участках губы на глубинах до 18 м, на валунных и смешанных грунтах. Видовое разнообразие достаточно велико: индекс Симпсона составляет 0,366, Шеннона – 0,771, видовое богатство – 19,0 видов/м². Сообщества монодоминантные, доля S. latissima составляет в среднем 63,38 ± 17,97%. В западной части губы (станции 6-8) субдоминантом является A. esculenta, на станциях 11 и 16 в центре губы – D. aculeata, на станции 12 – L. digitata. Велико разнообразие красных и зеленых, но существенную биомассу формируют только O. dentata (на некоторых станциях) и C. melagonium на станции 11.
2. Фитоценоз L. hyperborea + L. digitata + A. esculenta + S. latissima + D. aculeata с многоярусной вертикальной структурой и обильными синузиями эпифитов стволика и ризоидов ламинарий. Располагается в слабозащищенных участках губы на валунных грунтах на глубинах от 1 до 10 м. Видовое разнообразие наиболее высокое: индекс Симпсона – 0,17, Шеннона – 1,466, но видовое богатство только 19,3 вида/м². Сообщества полидоминантные, доля доминанта 28,31 ± 2,52%. Основная биомасса приходится на L. digitata, L. hyperborea, A. esculenta, S. latissima, D. aculeata, D. viridis, доля других видов невелика.
3. Фитоценоз S. latissima + бурые нитчатки расположен на песчаных грунтах с битой ракушей и отдельными разреженными валунами. Ассоциация отличается от остальных высоким разнообразием бурых водорослей второго яруса, из которых по массе преобладает Chorda filum (табл. 1 a, b, c). По составу доминантов и видовому разнообразию сходен с фитоценозом S. latissima + A. esculenta + D. aculeata: индекс Симпсона – 0,443, Шеннона – 0,838, видовое богатство – 18,3 вида/м².
4. Фитоценоз S. latissima без четкого выделения ярусов. К сопутствующим видам можно отнести только D. aculeata и U. obscura, но их биомасса менее 10% от доминанта. Это сообщества с существенно меньшим, чем в предыдущих группах, видовым богатством (табл. 1 a, b, c), хотя индексы видового разнообразия достоверно не отличаются из-за сравнительно высокой выравненности видов по обилию (табл. 3). Произрастает на илистых грунтах при слабом движении воды в защищенной части губы на глубинах 6-10 м.
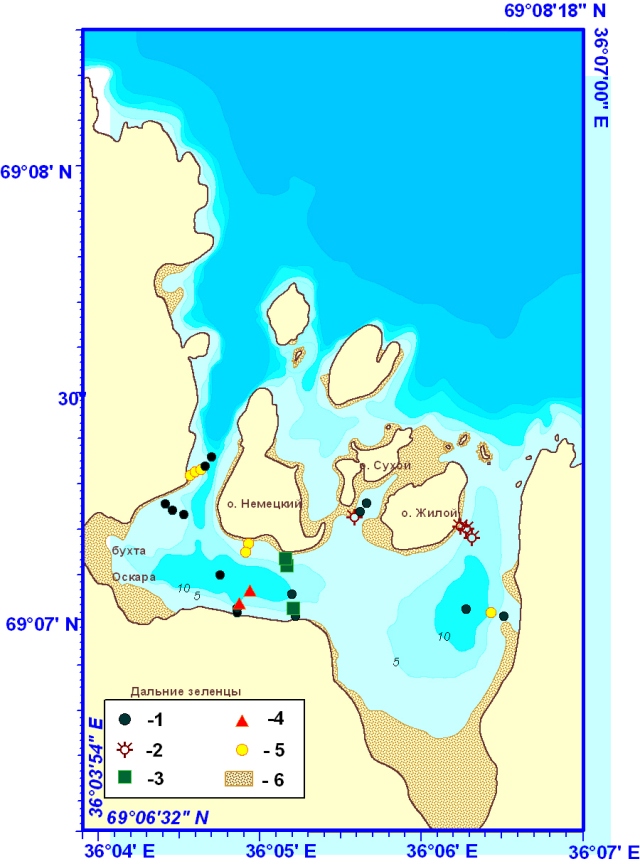
Рис. 3. Состав растительности в губе Зеленецкая. Цифрами обозначены фитоценозы:
1 - S. latissima + A. esculenta + D. aculeata; 2 – L. hyperborea + L. digitata + A. esculenta + S. latissima + D. aculeata; 3 - S. latissima + бурые нитчатки; 4 – S. latissima; 5 – станции с неопределенными пока фитоценозами; 6 – литораль
Выделение самостоятельного фитоценоза по кластеру из станций 3 и 4 в данном случае не представляется возможным вследствие малого количества имеющихся данных. В указанных фитоценозах видовое богатство низкое (8 и 6 видов/м²), но выравненность видов по обилию, выраженная индексами видового разнообразия, достаточно высокая (табл. 2). Относительно обильны P. rubens и O. dentata, зелёные водоросли отсутствуют. Биомасса доминантов низкая: на ст. 3 доминантами являются L. digitata и A. esculenta, что позволяет рассматривать ее как переходную от олигодоминантных фитоценозов станций 1 и 2 к обедненному сообществу D. aculeata на станции 4. Поскольку границы между фитоценозами достаточно размытые, вероятно, что ст. 3 была заложена в пограничной зоне. Также не вполне ясна принадлежность станций 1 и 2. Поэтому фитоценозы западного пролива губы Зеленецкая, в которых доминирует L. digitata, требуют более тщательного исследования.
Таким образом, в устьевой части губы до глубины 10 м на валунных грунтах растительность слагает фитоценоз L. hyperborea + L. digitata + A. esculenta + S. latissima + D. aculeata. В защищенной части губы на смешанных и валунных грунтах преобладает фитоценоз S. latissima + A. esculenta + D. aculeata, на илистых грунтах – S. latissima. В центральной части губы на грунтах, состоящих из песка с битой ракушей и отдельными валунами, растительность сложена фитоценозом S. latissima + бурые нитчатки. Сообщества L. digitata в западном проливе и S. dermatodea в восточном котле губы Зеленецкая пока не могут быть отнесены к определенному фитоценозу.
Сходные фитоценозы были описаны в губах Ивановская и Дроздовка Восточного Мурмана (Малавенда, Метельский, 2013). В слабозащищенной части губы Дроздовка (фиордового типа) растительность сложена фитоценозом S. latissima + A. esculenta, в куту губы – S. latissima + D. aculeata. В устьевой части губы Ивановская на глубине 5 м обнаружен фитоценоз L. digitata + A. esculenta, на 10 м – S. latissima + A. esculenta. В ковше и лагуне, отделенных от моря мелководными перемычками, выявлен фитоценоз L. digitata + S. latissima. В целом, при сходных условиях, фитоценозы в трех сравниваемых губах Восточного Мурмана являются аналогичными. Растительность губы фиордового типа Ярнышная (Шошина, Аверинцева, 1994) частично сложена сходными фитоценозами. В сублиторали защищенной от прибоя части губы преобладают фитоценозы, которые авторы отнесли в ассоциацию S. latissima + D. aculeata + бурые нитчатки. Приурочена данная ассоциация к смешанным грунтам (валун, гравий, песок и ил), заросли мозаичные, проективное покрытие дна водорослями сильно варьирует в зависимости от типа грунта. На глубинах 4-6 м в средней и 5-14 м в открытой части губы выделена асс. S. latissima + A. esculenta, массовыми видами в которой также являются D. aculeata и L. digitata. На смешанных и валунных грунтах широко распространена ассоциация L. digitata + S. latissima + D. aculeata, одним из массовых видов в которой является A. esculenta. В устье губы фитоценозы сложены асс. L. digitata + Ptilota plumosa (вероятно, соответствует Ptilota gunneri и P. serrata в описании К.Л. Виноградовой, 2011). На 10-20 м на илистых грунтах повсеместно отмечены крайне разреженные сообщества S. latissima и D. aculeata. Таким образом, видно, что распределение доминирующих видов аналогично в обеих губах. Приведенных в статье (Шошина, Аверинцева, 1994) данных недостаточно, чтобы однозначно соотнести выделенные сообщества, но по имеющейся характеристике фитоценоз S. latissima + D. aculeata + бурые нитчатки близок к описанному нами в губе Зеленецкая. Можно соотнести фитоценоз L. digitata + S. latissima + D. aculeata с выделенным нами L. hyperborea + L. digitata + A. esculenta + S. latissima + D. aculeata. Фитоценоз L. digitata + L. hyperborea + A. esculenta в губе Зеленецкая близок по составу и структуре с выделенной ранее ассоциации L. hyperborea + A. esculenta + L. digitata + S. latissima (Kain, 1962; 1963).
Сравнение биомассы и плотности популяции массовых видов водорослей во всех районах Восточного Мурмана показало их достаточную однородность (Блинова, 1969; 2007; Возжинская, 1996; Пропп, 1971; Кузнецов, Шошина, 2003). Анализ результатов исследований, проведенных в губах Зеленецкая (данная работа), Ярнышная (Шошина, Аверинцева, 1994), Ивановская и Дроздовка (Малавенда, Метельский, 2013) также позволяет предположить наличие общих закономерностей видового состава макроводорослей и распределения фитоценозов в заливах Восточного Мурмана.
Заключение
В результате анализа видовой структуры и пространственного распределения сублиторальных фитоценозов защищенной губы Зеленецкая Мурманского побережья Баренцева моря было обнаружено 67 видов макроводорослей и выделено 4 фитоценоза: L. hyperborea + L. digitata + A. esculenta + S. latissima + D. aculeata, S. latissima + A. esculenta + D. aculeata, S. latissima и S. latissima + бурые нитчатки. Мозаичность растительности свойственна фитоценозам, произрастающим в защищенных участках губы на илисто-песчаных грунтах. Вертикальная структура более четко выражена в фитоценозах с ярко выраженным доминантом. Описанная в работе видовая структура фитоценозов и их приуроченность к определенным местам обитания характерна также для открытых губ и губ фиордового типа Восточного Мурмана.
Полученные данные вносят вклад в понимание механизмов функционирования прибрежных биоценозов и имеют большое значение при выполнении экологических экспертиз в данном секторе Арктики.
Благодарности
Выражаем глубокую и искреннюю благодарность с.н.с. лаборатории альгологии БИН РАН к.б.н. Т.А.Михайловой за консультации при идентификации проб и обсуждении работы, а также сотрудникам лаборатории альгологии ММБИ КНЦ РАН м.н.с. А.А. Метельскому за помощь при обработке проб и с.н.с. к.б.н. И.В. Рыжик за помощь при отборе проб и выполнении подводной фотосъемки.
Работа выполнена при частичной финансовой поддержке РФФИ (грант № 11-04-90706 моб_ст).
Список литературы
1. Блинова Е.И. Вертикальное распределение и количественный учет макрофитов Айновских остовов (Баренцево море) / Распределение и состав промысловых водорослей Баренцева моря. // Тр. ММБИ. Вып. 8 (12). М.-Л.: Наука, 1965 а. С. 41-55.
2. Блинова Е.И. Распределение и запасы промысловых водорослей Мурманского побережья Баренцева моря: Автореф. дис. канд. биол. наук. Москва, 1965 б. 27 с.
3. Блинова Е.И. Водоросли-макрофиты и травы морей Европейской части России (флора, распространение, биология. запасы, марикультура). М.: Изд-во ВНИРО, 2007. 114 с.
4. Василевич В.И. Статистические методы в геоботанике. Л.: Наука, 1969. 232 с.
5. Виноградова К.Л. Виды Ceramium (Ceramiaceae, Rhodophyta) в северных морях России // Ботанич. журн. 2005. Т. 90. № 6. С. 884-890.
6. Виноградова К.Л. Виды Porphyra (Bangiales, Rhodophyta) в северных морях России // Ботанич. журн. 2007. Т. 92. № 4. С. 532-543.
7. Виноградова К.Л. Порядок Ceramiales (Rhodophyta) во флоре Северного Ледовитого океана // Ботанич. журн. 2011. Т. 96. № 6. С. 681-695.
8. Виноградова К.Л. Роды Chaetomorpha Kütz. и Rhisoclonium Kütz. (Siphonocladales) в северных морях СССР // Новости систематики низших растений. 1986. Т.23. С. 13-25.
9. Виноградова К.Л. Ульвовые водоросли (Chlorophyta) морей СССР. Л.: Наука, 1974. 166 с.
10. Возжинская В.Б. Макрофитобентос высокобореальных (холодноводно-умеренных) морей России: Автореф. дис. док. биол. наук. - Москва, 1996. - 46 с.
11. Завалко С.Е., Шошина Е.В. Многоуровневая морфофизиологическая оценка состояния фукусовых водорослей в условиях антропогенного загрязнения (Кольский залив, Баренцево море) // Вестник МГТУ. 2008. Т. 11, № 3. С. 423-431.
12. Зинова А.Д. Определитель бурых водорослей северных морей СССР. М.-Л.: Изд-во АН СССР, 1953. 225 с.
13. Зинова А.Д. Определитель красных водорослей северных морей СССР. М.-Л.: Изд-во АН СССР, 1955. 220 с.
14. Кузнецов Л.Л., Шошина Е.В. Фитоценозы Баренцева моря (физиологические и структурные характеристики). Апатиты: Изд-во КНЦ РАН, 2003. 308 с.
15. Лебедева Н.В., Дроздов Н.Н., Криволуцкий Д.А. Биологическое разнообразие. М.: Владос, 2004. 432 с.
16. Лоция Баренцева моря. Часть 2. Адм. № 1112. От реки Ворьема до пролива Карские Ворота и западные берега острова Новая Земля // СПб: ГУН и О. 1995. 464 с.
17. Макаров В.Н., Макаров М.В., Шошина Е.В. Современное состояние зарослей ламинарии сахаристой Баренцева моря, методы прогнозирования запасов и паспортизация ее коммерческой ценности. Препринт. Апатиты, 1997. 44 с.
18. Малавенда С.В., Метельский А.А. Ассоциации ламинариевых водорослей губ Ивановская и Дроздовка // Вестник МГТУ. 2013. Т. 16. № 3. С. 493-500.
19. Пельтихина Т.С. Ламинариевые водоросли Баренцева моря и их рациональное использование. Мурманск: Изд-во ПИНРО, 2005. 123 с.
20. Перестенко Л.П. Род Aсrosiphonia J.Ag. на Мурманском побережье (Баренцево море) // Новости систематики низших растений. 1965а. Т.2. С.50-64.
21. Пригоровский Б.Г. Фауна мягких грунтов литорали губы Дальне-Зеленецкой / Тр. Мурм. биол.ст. Т.1. М.-Л.: Изд-во АН СССР, 1948. С. 146-155.
22. Промысловые и перспективные для использования водоросли и беспозвоночные Баренцева и Белого морей. Апатиты: Изд-во КНЦ РАН, 1998. 628 с.
23. Пропп М.В. Экология прибрежных донных сообществ Мурманского побережья Баренцева моря. Л.: Наука, 1971. 128 с.
24. Сорокин А.Л. Ландшафты шельфа Кольского полуострова: геолого-геоморфологические основы формирования. Мурманск: Кн. изд-во, 1987. 128 с.
25. Суханов В.В., Жуков В.Е. Закономерности в изменчивости видовой структуры прибрежного сообщества водорослей-макрофитов: модельный анализ // Журн. общ. биол. 2003. Т. 64. № 3. С. 248-262.
26. Тиховская З.П. Видовой состав морских водорослей в районе Мурманской биологической станции. / Тр. Мурм. биол.ст. Т.1. М.-Л.: Изд-во АН СССР, 1948. 189-191.
27. Ушаков П.В. Мурманская биологическая станция Академии Наук СССР в губе Дальне-Зеленецкой и ее первые научные работы / Тр. Мурм. биол.ст. Т.1. М.-Л.: Изд-во АН СССР, 1948. С. 10-33.
28. Шошина Е.В., Аверинцева С.Г. Распределение ассоциаций водорослей, видовой состав и биомасса в губе Ярнышной Баренцева моря / Гидробиологические исследования в заливах и бухтах северных морей России. Апатиты: Изд-во КНЦ, 1994. С. 38-61.
29. Field J.G., Clarce K.R., Warwick R.M. A practical strategy for analyzing multispecies distribution pattern // Mar. Ecol. Prog. Ser. 1982. V. 8. P. 37-52.
30. Guiry M.D., Guiry G.M. AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org; searched on 28 January 2014.
31. Hammer Ø., Harper D. A. T., Ryan P. D. PAST - PAlaeontological STatistics, ver. 1.87. – 2008.
32. Kain J. M. Aspects of the biology of Laminaria hyperborea I. Vertical distribution // J. Mar. Biol. Ass. U.K. 1962. V. 42. №. 2. P. 377-385.
33. Kain J.M. Aspect of the biology of Laminaria hyperborea. II. Age, weight and length // J. mar. boil. Ass. U. K. 1963. Vol. 43. P. 129-151.
опубликовано - октябрь 2014 г.
The structure of sublittoral phytocenoses in wave-protected bay Zelenetskaya, Murman coast, Barents sea
Svetlana V. Malavenda, Mikhail V. Makarov
The analysis of the spatial distribution and species composition of subtidal phytocenoses in wave-protected Zelenetskaya bay (Murman coast of the Barents Sea) was carried out. There were 4 association allocated in the bay: L. hyperborea + L. digitata + A. esculenta + S. latissima + D. aculeata, S. latissima + A. esculenta + D. aculeata, S. latissima и S. latissima + brown filamentous algae. Described specific structure of phytocenoses and their association with specific habitats is typical also for open bays and fjord-type bays of Eastern Murman.
Key words: phytocenoses; phytocenoses’ structure; species diversity; subtidel zone; Murman coast; Barents Sea.
Об авторах
Малавенда Светлана Владимировна - Malavenda Svetlana V.
кандидат биологических наук
ММБИ КНЦ РАН (Murmansk marine biological institute KSC RAS)
malavenda@yandex.ru
Макаров Михаил Владимирович - Makarov Mikhail V.
доктор биологических наук
ММБИ КНЦ РАН (Murmansk marine biological institute KSC RAS)
science@mmbi.info
Корреспондентский адрес: Россия, 183010, ул. Владимирская, 17, Мурманск, ММБИ КНЦ РАН; тел. 8-8152-250696, факс 8-8152-253994.
ССЫЛКА НА СТАТЬЮ:
Малавенда С.В., Макаров М.В. Структура сублиторальных фитоценозов защищенной губы Зеленецкая Мурманского побережья Баренцева моря // Вопросы современной альгологии. 2014. № 1 (5). URL: http://algology.ru/472
При перепечатке ссылка на сайт обязательна
На ГЛАВНУЮ

КОНТАКТЫ
Email: info@algology.ru
Изготовление интернет сайта
5Dmedia
ЛИЦЕНЗИЯ
Эл N ФС 77-22222 от 01 ноября 2005г.
ISSN 2311-0147