|
|
|
О выделении ассоциаций морских донных макрофитов северо-западной части Татарского пролива
Дуленин А.А. Alexander A. Dulenin
Хабаровский филиал ФГБНУ «Всероссийский научно-исследовательский институт
УДК 574.58
На основании анализа материалов альгологических съемок в северо-западной части Татарского пролива разработана методика выделения сообществ донных макрофитов на основании доминантно-физиономического подхода. Доминанты ассоциаций установлены по превышению заранее установленных порогов обилия. Каждая ассоциация выделена и поименована по единственному наиболее обильному доминанту главного яруса. При описании ассоциаций использованы два показателя обилия – проективное покрытие (приоритетный) и удельная биомасса (дополнительный). Использование количественных показателей обилия делает полученные характеристики проверяемыми, сравнимыми и доступными для статистической обработки. Предложенная методика позволяет однозначно опознавать и картировать ассоциации визуально. Она учитывает особенности проведения альгологических исследований при выполнении съемок вдоль протяженных побережий в условиях дальневосточных и северных морей России. Ключевые слова: Татарский пролив; сублитораль; выделение сообществ макрофитов
Введение Важнейшей задачей ресурсных и прикладных экологических исследований является изучение закономерностей распределения донной морской растительности, в частности выделение, описание и картирование ее сообществ, к которым пространственно приурочены запасы промысловых видов растительности и поселения экологически и хозяйственно значимых растений и животных. Между тем, сообщества сублиторальной растительности морей России в целом изучены слабо и неравномерно, а усилия, направляемые на изучение растительности морей, весьма малы. Например, из 152 публикаций конференции «Гидроботаника 2010» (2010), морской растительности посвящено только 5 статей, т.е. 3%. Наиболее подробно описана растительность Черного и Азовского морей (Калугина-Гутник, 1975; Громов, 1998; Афанасьев, Рубан, 2013; и др.). Описана растительность Белого моря (Возжинская, 1986). В дальневосточных морях была подробно исследована сублиторальная растительность лишь нескольких больших регионов: Курильских островов (Гусарова, 1975), южной части Сахалина (Петров, Поздеев, 1992), северо-западной части Татарского пролива (Дуленин, 2008). Большинство исследований донной морской растительности Дальнего Востока выполнено на небольших участках в южном Приморье. Список этих публикаций приводят Т.Л. Калита и А.В. Скрипцова (2018). Однако донные растительные сообщества этих регионов, как правило, были описаны либо умозрительно, либо без указания четких количественных критериев их выделения. Вследствие этого полученные результаты трудно проверять и сравнивать, они не всегда могут быть повторены. Четкие количественные критерии выделения сообществ и оценок доминирования приводятся лишь в ряде работ последних лет, выполненных в южном Приморье (Скрипцова и др., 2013; 2018; Калита, Скрипцова, 2014; 2018). Однако в них для оценки доминирования видов используются только показатели биомассы. Между тем, важнейшей количественной характеристикой видов сообщества, отражающей успех растений в конкуренции за субстрат (Набивайло, Титлянов, 2006), является проективное покрытие (Миркин и др., 1989), которое авторы упомянутых выше работ не учитывали. Кроме того, при определении степени доминирования они использовали относительные доли обилия видов в сообществе, что приводит к артефактам в оценках доминирования при слабо развитом растительном покрове. Также они применяли неравномерную шкалу обилия, что не всегда удобно при статистической обработке данных. Авторы использовали избранную ими методику лишь для описания отдельных сообществ мелководий нескольких локальных акваторий. В то же время, удовлетворительного методического подхода, который позволял бы на основании стандартизированных количественных критериев описывать сублиторальную растительность больших морских районов во время учетных съемок, к настоящему времени, по-видимому, не выработано. Вообще, исследования морской растительности в целом строятся на методических наработках, полученных в наземной геоботанике, а особенности растительного покрова донных макрофитов учитываются относительно редко (Миркин и др., 1989; Блинова и др., 2005). Обоснования методологических основ описания сообществ и их классификации в связи с особенностями состава, структуры и исследований донной морской растительности, судя по всему, до сих пор сделано не было. Работы, в которых давалась бы синтаксономическая классификация донной растительности дальневосточных морей России, до сих пор отсутствуют. В настоящее время в России и в мире преобладает флоро-ценотический подход к описанию растительности, основанный на геоботанической системе Ж. Браун-Бланке (Миркин, Наумова, 2009) с принципами и правилами, регулируемыми Международным кодексом фитосоциологической номенклатуры (Вебер и др., 2005). Однако, характеристики растительного покрова различных участков Земли столь различны, а условия его изучения так многообразны, что нужно признать если не принципиальную невозможность, то во всяком случае практическую нецелесообразность использования единственного методологического подхода для всех случаев. Показательно, что даже при использовании флоро-ценотического подхода «Около 5/6 ассоциаций выделено по доминантам, которые выступают единственными характерными видами ассоциаций» (Василевич, 2010). В общем случае, подходы к описанию и классификации растительности должны обеспечивать принципиальную возможность верифицируемости (проверяемости) и фальсифицируемости (опровергаемости) научного исследования (Popper, 2002). Выбор методологических основ исследования должен диктоваться основными свойствами его предмета, в нашем случае – донной морской растительности, а также задачами и реальными возможностями исследования. В целом, избранный методологический подход должен обеспечивать получение адекватной характеристики растительного покрова при минимуме затрат труда и времени. Исходя из этих соображений, использование флоро-ценотического подхода оптимально для морей, где доминирование отдельных видов не выражено (Афанасьев, Абдуллин, 2013). Однако, в тех районах морей, где хорошо проявляется доминирование, эколого-фитоценотический (или доминантно-физиономический) подход представляется более продуктивным, в частности, для маловидовых, моно- и олигодоминантных сообществ (Афанасьев, Рубан, 2013). Далее, Международный кодекс фитосоциологической номенклатуры (Вебер и др., 2005), рекомендует сопровождать публикации полными первичными данными геоботанических описаний. Между тем, такие сведения, особенно в отношении сообществ, виды которых представляют промысловую ценность, предназначены для служебного пользования, составляют коммерческую тайну, и поэтому не допускаются к опубликованию, что не позволяет использовать этот принцип флоро-фитоценотического подхода для описания таких сообществ. Кроме того, флоро-фитоценотический подход подразумевает как можно более полное описание видового состава каждой ассоциации. Однако практика показывает, что во время ограниченных по времени водолазных съемок вдоль протяженных берегов дальневосточных и северных морей России, в тяжелых условиях водолазных погружений (низкие температуры воды, течения, плохая видимость), технические возможности для выполнения флористических сборов с необходимой тщательностью представляются редко, тем более, что большую часть материалов собирают водолазы-небиологи. В таких случаях практически целесообразным оказывается выделение сообществ по их физиономическому облику (Taniguti, 1962). Исходя из изложенного, автор для описания и классификации растительности использовал доминантно-физиономический подход. Автор в настоящей работе попытался решить две основные задачи: 1) выработать методику описания олигодоминантных сублиторальных морских сообществ протяженных морских районов на основании четко обозначенных количественных критериев: проективного покрытия (ПП) и удельной биомассы (УБМ), которая позволяла бы с наименьшими трудозатратами получать сравнимые и проверяемые результаты; 2) выполнить синтаксономическую классификацию сублиторальной растительности протяженного морского района и составить ее продромус.
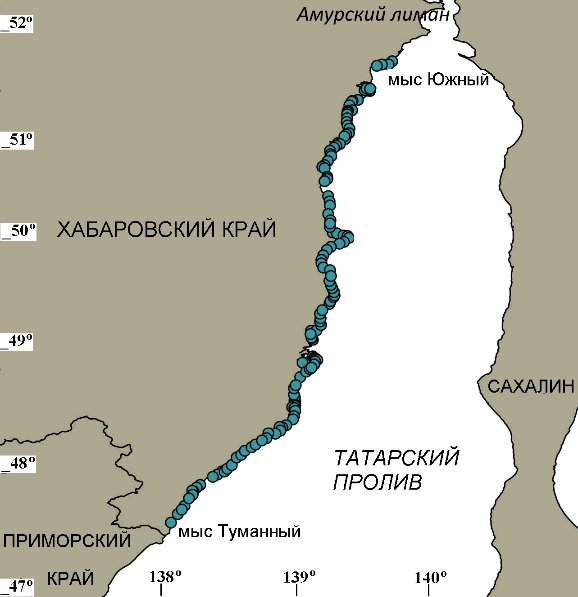
Материалы и методы В качестве модельного района для исследования была выбрана северо-западная часть Татарского пролива, протяженного на 600 км с юга на север, что позволяет описывать растительность на региональном инфраценотическом уровне (Миркин и др., 2001). Материалы собраны в экспедициях «ХабаровскНИРО» (ранее «ХфТИНРО») с 1999 по 2020 гг. в районе от мыса Туманного на юге (47°24' N, 139°03' E) до мыса Южного на севере (51°40' N, 141°06' E), т.е. в пределах Хабаровского края (рис. 1, табл. 1). Во время НИР выполняли водолазные съемки, подводные наблюдения с помощью привязных видеокамер и телеуправляемых необитаемых подводных аппаратов (ТНПА), наблюдения с поверхности. Кроме того, выполняли аэрофотосъемки при помощи беспилотных летательных аппаратов (БПЛА). Всего было проведено 13 экспедиций, во время которых выполнено 2049 станций съемок. Во всех съемках автор принимал непосредственное участие. Не менее 600 погружений (гидробиологических водолазных станций) автор выполнил лично. Основной объем материалов (711 станций водолазной съемки) собран во время экспедиции на НИС «Убежденный» в июле 2010 г. Вдоль всего района сборы проведены на 150 водолазных разрезах, выполненных перпендикулярно берегу на глубинах от 2 до 20 м (местами от 0 до 22 м). Водолазные станции на разрезах были расположены в пределах основных ценотических поясов. При выполнении станций руководствовались типическим подходом (Блинова и др., 2005). Расстояние между разрезами составляло от 0,5 до 6 миль (в среднем 2 мили), в зависимости от однородности берегов и структуры растительности. Удельную биомассу (УБМ, кг/м2) учитывали путем отбора с площадок в 1 м2 для относительно редко расположенных растений и в 0,25 м2 с последующим пересчетом на 1 м2 – для плотно расположенных. ПП (долю покрытия дна видом) определяли глазомерным способом без использования инструментальных методов путем осмотра 10–100 м2 дна, в зависимости от характера поселений и видимости. В условиях плохой видимости ПП не учитывали. При ПП от 10 до 100% использована равномерная шкала ПП (Миркин и др., 1989) с шагом в 10%. При меньшем ПП его указывали с точностью до 1%.
Одна гидробиологическая станция соответствовала одному гидроботаническому описанию. При выделении сообществ автор руководствовался прагматическим подходом, исходя из принципа редукционизма (Миркин и др., 1989), в соответствии с которыми выполнялся лишь необходимый минимум классификационных построений, необходимый для однозначного опознания ассоциаций и синтаксонов высшего порядка. Для выделения ассоциаций оценки ПП считались приоритетными (Дуленин, 2020), УБМ – дополнительными. Для каждой ассоциации выделяли 3 категории видов по степени доминирования – доминанты, субдоминанты и сопутствующие. Ассоциации опознавали по главному доминанту (ГД), принадлежащему к «доминирующему ярусу наибольшей высоты» (Вебер и др., 2005), т.е. наиболее простым и логичным для их визуального выявления способом. В качестве доминанта каждого яруса указывался наиболее обильный в этом ярусе вид при условии достижения порогов доминирования (Баканов, 1987), условно принятых ранее для растительности района (Дуленин, 2020): по ПП – 0,3, по УБМ – 1 кг/м2. Пороги доминирования были использованы, чтобы избежать артефактов описания растительности разреженных поселений, когда в качестве доминантов могут быть указаны заведомо малообильные виды, не формирующие растительного покрова. Виды, максимальное обилие которых было меньше, чем у доминантов, однако не ниже пороговых, обозначены в качестве субдоминантов. Виды с максимальными показателями обилия ниже пороговых описаны в качестве сопутствующих. Максимальную ценотическую роль видов в пределах ассоциации указывали в связи с тем, что на отдельных станциях ценотическая роль видов могла меняться от сопутствующих до доминантов. В качестве принадлежащих к одной ассоциации описывались условно однородные участки растительности (Миркин и др., 2001) множества (не менее трех) гидроботанических описаний с одним и тем же ГД. Хотя выделение ассоциаций на основании одиночных описаний допустимо, такие ассоциации рассматриваются как сомнительные (Вебер и др., 2005), поэтому данные, полученые на 1–2 станциях исключены из анализа. Процедура описания ассоциаций была следующей. Основой для них служили стандартные электронные таблицы с описаниями гидробиологических станций (см. табл. 1), содержащие сведения об отмеченных на станции видах и показателях их обилия. Для выделения ассоциаций в эти таблицы были добавлены столбцы (Д1–Д4), в которые для каждой станции вписывали доминант каждого яруса, начиная с первого (верхнего) по четвертый (нижний). Кроме того, в таблицу был добавлен столбец с указанием яруса, к которому относится каждый вид (первый – крупные бурые водоросли и морские травы с высотой растительного покрова от 0,4 до 4 м, второй – пластинчатые и кустистые водоросли высотой от 5 до 40 см, третий – кустистые водоросли высотой до 5 см, и четвертый – корковые водоросли). Сначала из электронной таблицы с общим описанием съемки отфильтровывали станции, на которых доминировали растения первого яруса. Каждый такой малый массив с доминированием одного вида первого яруса вырезали из общей электронной таблицы и помещали на отдельный лист. Такие массивы служили основой для описания отдельных ассоциаций и статистической обработки количественных данных. Для этого строили сводную таблицу, где в названиях строк приводили номера станций, в названиях столбцов – виды, а в теле таблицы – показатели обилия (ПП и УБМ) видов сообщества на каждой станции. Эту таблицу использовали для дальнейшей статистической обработки. Виды выделенных ассоциаций ранжировали по убыванию средних значений обилия, начиная с верхнего яруса до нижнего, указывая сначала доминант, потом субдоминанты, и наконец, сопутствующие виды. В первую очередь виды ранжировали по ПП, во вторую – по УБМ. Для каждого вида приводили количество описаний (т.е. гидробиологических станций), на которых этот вид встречался в пределах ассоциации, а также минимальное, максимальное, среднее и медианное значения обилия отдельно для ПП и УБМ и частоту встречаемости вида в сообществе (ЧВС). Ниже списка видов каждого яруса приводили суммарные данные по его обилию. Ниже списка всех видов ассоциации указывали суммарные показатели обилия этой ассоциации. Результаты обработки представляли в виде итоговой таблицы (см. табл. 2). Исключив из таблицы первичных данных съемки все данные об ассоциациях первого яруса, всю последовательность действий выполняли для ассоциаций, опознаваемых по главному доминанту своих более низко расположенных верхних ярусов, – последовательно второго, третьего и четвертого. Для каждой ассоциации вычисляли максимальное число ярусов. Самостоятельные ярусы в каждой ассоциации выделяли только в том случае, если суммарное максимальное обилие его видов формировало выраженный растительный покров с ПП≥0,3 и УБМ≥1 кг/м2. Если суммарное обилие растений, принадлежащих к определенному ярусу, не достигало порогового, ограничивались перечислением видов с указанием их обилия без выделения самостоятельного яруса. Для всех ассоциаций были определены предельные глубины и широты, участки распространения, частота встречаемости (ЧВ, % от всех станций съемки, на которых были отмечены растения). Поскольку оконтуривания границ отдельных фитоценозов не проводилось, ЧВ показывала относительную долю площади описываемой ассоциации от общей площади растительности. Для каждой ассоциации указывали видовое богатство в форме ∑, m(min-max), где ∑ – видовое богатство ценофлоры ассоциации, а m(min-max) – среднее, минимальное и максимальное видовое богатство на отдельных гидробиологических станциях. Для синтаксономической классификации использован подход Е.И. Вейсберга (2010), совмещающего использование доминантного и флористического методов классификации, при котором доминанты ассоциаций обозначаются в качестве их характерных, т.е. «предпочитающих тот или иной синтаксон» (Миркин и др., 1989) видов. Названия ассоциаций образовывали из видового названия доминантов, используя рекомендации Международного кодекса фитосоциологической номенклатуры (Вебер и др., 2005). При их наименовании было принято, что в названии будут указываться только доминанты либо субдоминанты нижних ярусов с относительно высокой степенью постоянства, т.е. превышающие порог доминирования в большей части гидроботанических описаний (>50%). В иных случаях субдоминанты и доминанты нижних ярусов было принято указывались в качестве таковых только в описательной таблице сообщества. Ассоциации с главными доминантами, принадлежащими к одному роду, объединяли в союзы. В синтаксоны ранга порядка объединяли союзы с главными доминантами своих ассоциаций, таксономически принадлежащими к одному порядку, в синтаксоны ранга класса – порядки с главными доминантами, таксономически принадлежащими к одному классу. Результаты обобщения данных сведены в итоговую таблицу (см. табл. 3), содержащую качественные и количественные характеристики каждой ассоциации. Описанные ранее разреженные поселения (Дуленин, 2008, 2019), не образующие выраженного растительного покрова, где растения не формируют между собой структурных связей, а суммарное обилие не достигает порогов доминирования, не рассматривались. Из-за невозможности опубликования в соответствии с рекомендациями Международного кодекса фитосоциологической номенклатуры (Вебер и др., 2005) первичных гидроботанических описаний, представляющих коммерческую ценность, таблицы 2 и 3 содержат вместо таких описаний обобщенные статические данные. Географические названия приведены по Лоции Японского моря (Лоция …, 2003). Табличные данные обрабатывали в MS Excel, статистические вычисления выполняли в свободно распространяемом специализированном пакете PAST 3 (Hammer, 2018). Виды идентифицированы по определителям (Перестенко, 1994; Клочкова, 1996). Названия таксонов приведены по AlgaeBase (Guiry, Guiry, 2021). Данные по корковым кораллиновым водорослям (ККВ) приведены без разделения по видам в связи со сложностью отбора их проб в полевых условиях.
Результаты и обсуждение Практика показывает, что первым и необходимым условием выделения и описания сообществ, обеспечивающим проверяемость и воспроизводимость результатов, является стандартизированная система записи первичных данных, обеспечивающая возможность их дальнейшей автоматизированной обработки. Имеются случаи, когда несоблюдение этого условия затрудняло анализ результатов съемок, а то и вовсе делало полученные данные непригодными для использования. Опыт рассмотрения данных гидробиологических съемок (по крайней мере, в отношении донной морской растительности) свидетельствует, что несмотря на кажущуюся простоту, система табличной записи первичных данных, которая позволяла бы удовлетворительным образом их обрабатывать, вовсе не является самоочевидной. В связи с этим, прежде всего автор находит нужным привести пример оформления такой таблицы (табл. 1) с некоторыми необходимыми пояснениями. Первичные данные о растительности, оформленные в виде стандартизированных электронных таблиц, позволили встроенными средствами фильтров, сводных таблиц, а также внешними средствами статистической обработки данных рассортировать и обобщить качественные и количественные сведения о растительном покрове. Важно, чтобы описание каждой станции содержало столько строк, сколько на ней обнаружено видов. Общие описательные данные (глубина, грунт и т.п.) каждой станции являются постоянными и повторяются в каждой строке, а данные о видах и их обилии – переменными, для них в каждой строке вносятся собственные данные. Такие таблицы позволяют выделять и описывать отдельные растительные ассоциации и синтаксоны более высокого порядка, анализировать особенности их состава, структуры и распространения. Таблицы можно модифицировать и дополнять новыми данными. Например, включение в таблицу 1 столбца с датой сбора позволяет собирать и анализировать данные о сезонных аспектах фитоценозов, о температуре воды – отслеживать изменения обилия и видового состава, связанные с прогревом вод, и т.п. После выделения синтаксонов, их названия можно указать в отдельных столбцах (табл. 1, серая заливка). Необходимо, чтобы все данные об обилии, в том числе глазомерные ранговые, были записаны в числовой форме, с использованием равномерных шкал. Иначе сопоставление различных показателей обилия (например, ПП и УБМ), как и статистическая обработка данных, будет осложнена или невозможна. Для записи и обработки такого рода данных вполне достаточно простых плоских электронных таблиц. В то же время анализ более сложных данных (например, включающих дополнительно размерно-массовые характеристики отдельных видов растений, данные об эпифитах, ассоциированном бентосе, и т.п.) может требовать использования реляционных баз данных. Результаты обработки первичных данных представлены в виде итоговых таблиц, характеризующих качественный состав, уровни доминирования, вертикальную структуру и показатели обилия. Их средние со стандартными ошибками характеризуют обилие ассоциаций с репрезентативным количеством описаний, медианы – обилие в малых выборках. Здесь для примера рассмотрена таблица с характеристикой ассоциации Ulvetum lactucae. Описание ассоциации начато с растений верхнего яруса, хотя растения этого яруса расположены редко и ни один вид во всех обработанных описаниях не превышает установленного порога доминирования по ПП (виды 1–4 по списку из табл. 2). Stephanocystis crassipes достигает максимальной УБМ в 1 кг/м2, однако поскольку ПП при этом не превышает 0,1, следует понимать, что эти растения расположены поодиночке, разреженно. Суммарное обилие растений I яруса меньше порога доминирования и не создает выраженного растительного покрова, поэтому следует говорить о том, что этот ярус в ассоциации не формируется. Впрочем, в данном конкретном случае вероятно, что это наблюдение не закономерность, а следствие недостатка статистики: ЧВ ассоциации мала, поэтому ее характеристика дана всего по четырем гидроботаническим описаниям. Следует полагать, что новые описания существенно дополнят сведения о ее структуре и составе.
Таблица 1. Фрагмент электронной таблицы с основными данными первичных гидроботанических описаний. Отфильтрованы станции с доминированием Ulva lactuca. Некоторые столбцы (дата, номер разреза, удаление от берега, способ отбора проб, биомасса вида на учетной площади, примечания) исключены из фрагмента для экономии места THE TABLE CAN BE SEEN BY CLICKING ON THE LINK
ТАБЛИЦА ОТКРЫВАЕТСЯ ПРИ НАЖАТИИ НА ССЫЛКУ THE TABLE CAN BE SEEN BY CLICKING ON THE LINK
Во II ярусе растительности располагается ГД ассоциации – Ulva lactuca. Ее УБМ меньше пороговой, однако при ПП 0,3–0,5 вид образует выраженный растительный покров, занимая на участках своего доминирования наибольшую долю поверхности субстрата. Несмотря на то, что массивные растения Codium fragile – следующего по обилию вида этого яруса – имели УБМ 2 кг/м2, их ПП при этом составляло лишь 0,1, и поэтому здесь вид рассматривается в качестве сопутствующего, как и прочие виды этого яруса – Neohypophyllum middendorfii и Tichocarpus crinitus, данные о ПП которых в пределах рассматриваемой ассоциации собрать не удалось. Субдоминантов II яруса (С2), ПП которых было бы выше порогового, но ниже, чем у ГД, не выявлено. В этом случае об их обилии позволяют судить показатели УБМ. Максимальные и средние суммарные показатели обилия растительности, относимой ко II ярусу, превышают пороговые и позволяют выделить самостоятельный II ярус в пределах описываемой ассоциации. Из растений, относимых к III ярусу, отмечена только Bossiella compressa с небольшим ПП. Доминантов (Д3) и субдоминантов (С3), принадлежащих к этому ярусу, нет. Указанное обстоятельство не позволяет выделить самостоятельный III ярус в рассматриваемой ассоциации. Показатели УБМ имеют ключевое значение для оценки промысловых ресурсов, прикладных экологических исследований и прочих хозяйственно значимых работ. Количество проб ПП и УБМ в итоговой таблице (см. табл. 2) могут не совпадать, что естественно в условиях ограниченной по времени съемки у побережий большой протяженности: при высокой мутности на некоторых станциях бывает невозможно оценить ПП, при сильном волнении или течении – УБМ. При этом вернуться на станцию для сборов после наступления благоприятных условий при проведении ограниченной во времени подвижной морской экспедиции обычно не представляется возможным. В реальной практике, при невозможности сбора данных по ПП из-за высокой мутности, можно ограничиться только данными об УБМ. Если же съемка выполнена без применения водолазной техники (например, с использованием подводной видеоаппаратуры) можно наоборот, ограничиться данными о ПП без получения сведений об УБМ. Кроме того, при необходимости таблицы можно расширить, дополнив их данными о плотности, во всяком случае для крупных многолетних растений (Agarum clathratum, представители Sargassaceae, и т.п.). Преимущество использования как минимум двух показателей обилия растений, достаточно дробных равномерных ранговых шкал (ПП) и указания конкретных величин (кг/м2, экз./м2) заключается в возможности получения хозяйственных характеристик сообщества – биомасс видов, занимаемой ими площади и т.п. Кроме того, для ряда видов, у которых хорошо выражена зависимость между различными показателями обилия, возможны пересчеты одного показателя в другой (Дуленин, 2016). Итак, предложенная форма таблиц позволяет представлять данные о качественном составе, вертикальной структуре, показателях обилия, степени доминирования и частоте встречаемости видов растительных сообществ. При формировании документов для служебного и коммерческого использования (отчетов о НИР, разнообразных рыбохозяйственных характеристик и т.п.), для сравнительного описания ассоциаций или синтаксонов более высокого порядка, возможно составление их синоптических таблиц (Миркин и др., 2001). Однако вместо использования неравномерных относительных шкал обилия (Браун-Бланке, Друде, и т.п.) целесообразно приводить количественные данные об обилии, как показано выше. Ни для одной из описанных ассоциаций не обнаружены доминанты и субдоминанты подчиненных ярусов, которые были бы надежно связаны с ГД. Частота доминирования доминантов нижних ярусов и субдоминантов не превышает 50%, что не позволяет говорить об их приуроченности к ассоциациям и использовать их для выделения и наименования ассоциаций. Разумно предположить, что такая особенность может быть закономерной для морей умеренных широт, где на отдельных участках в каждом растительном сообществе наибольшего успеха в конкуренции за ресурсы добивается вид, становящийся ГД при том, что как присутствие, так и достижение значительных показателей обилия прочими видами более или менее случайно. Выполнена синтаксономическая классификация и составлен предварительный продромус растительности района исследований. В районе исследования описано 18 ассоциаций сублиторальных донных макрофитов (табл. 3). Выделенные ассоциации отнесены к четырем классам, семи порядкам и четырнадцати союзам. Очевидно, что предложенный продромус, как и сведения о составе и структуре ассоциаций будут дополняться во время дальнейших исследований. Поскольку ассоциации выделены по ГД, от которого произведены их названия, это позволяет выполнить синтаксономическую классификацию, которая параллельна таксономической, где названия синтаксонов высшего порядка образованы от названий соответствующих таксонов. Интересно, что такая синтаксономия экологически оправдана – во всяком случае в пределах района исследований. Например, внутри порядка Laminaretalia в союз Saccarinetum включены ассоциации, образующие свои фитоценозы в сходных экологических условиях на твердых грунтах у открытых побережий, но сменяющих друг друга по градиенту широты (Saccharinetum japonicae – S. cichorioidae) или глубины (S. japonicae – S. sculperae). Ассоциации союза Saccharinetum сменяются (как правило, с глубиной) единственными для своих союзов ассоциациями, принадлежащими союзам Costaretum, Desmarestetum, Agaretum. На мелководьях твердых грунтов ассоциации порядка Laminaretalia сменяются ассоциациями порядка Sargassetalia. Союзы этого порядка с их ассоциациями сменяются по мере снижения гидродинамической нагрузки (Sargassion – Stephanicystion). При дальнейшем снижении гидродинамической нагрузки синтаксоны класса Phaeophycetea сменяются синтаксонами Zosteretea – на мягких грунтах это ассоциации, принадлежащие к пор. Zosteretalia, а на мелководьях с твердыми грунтами – единственная ассоциация пор. Phyllospadetalia. Построенная синтаксономическая классификация не требует обязательного указания авторов синтаксонов, поскольку их объем с очевидностью определяется ГД ассоциаций. В случаях, когда построение подобной синтаксономии возможно, она представляется более удобной, чем флористическая, где классификация строится на основе консенсуса мнений исследователей, что ведет к неопределенности, частым ревизиям синтаксонов, вызывает необходимость указания автора и года описания синтаксона, без чего установить его смысл и объем бывает затруднительно (Вебер и др., 2005). Ряд ассоциаций (например Sargassetum miyabeae, Zosteretum asiaticae и др.) распространены в районе исследований повсеместно, другие – лишь на нескольких ограниченных акваториях (Zosteretum marinae) или вдоль небольшого участка побережья (Ulvetum lactucae). Они формируются на глубинах от 1 до 22 м, при этом одни могут встречаться практически по всему диапазону глубин формирования выраженного растительного покрова (Saccharinetum japonicae), другие – лишь в узком диапазоне (Costaretum costatae, Zosteretum marinae). Видовое богатство ценофлор ассоциаций измненялось в широких пределах – от 1 вида для Zosteretum asiaticae до 31 вида для Cortexetum corallinae, при этом видовое богатство на отдельных станциях было довольно низким, не более 12 видов. Очевидно, что довольно низкие показатели видового богатства связаны прежде всего со спецификой сжатых по времени водолазных съемок у побережий большой протяженности, когда тщательное определение видового состава не является приоритетом съемки, а большую часть сборов делают водолазы-небиологи. ЧВ отдельных ассоциаций в пределах пояса сублиторальной растительности изменялась от 1% (Ulvetum lactucae, Saccharinetum sculperae) до 9–15% (Saccharinetum japonicae и Agaretum clathratae соответственно). Суммарная ЧВ выделенных растительных ассоциаций – 68%. Остальные 32% составили разреженные поселения и отдельные участки растительности с выраженным доминированием отдельных видов, на которые пришлось по 1-–2 гидроботанических описаний, не отнесенные ни к одной из ассоциаций. Выделенные ассоциации надежно опознаются визуально, могут быть картированы при помощи ТНПА, а на прозрачных мелководьях – и с помощью БПЛА, а их количественные характеристики пригодны для статистической обработки и разнообразных хозяйственных вычислений. В соответствии с редукционистским подходом к классификации растительности (Миркин и др., 1989), автор полагает, что выделение более дробных синтаксонов практически нецелесообразно. При описании ассоциаций полезно как можно более полное выявление их флористического состава (Вебер и др., 2005). Однако в практике хозяйственно-значимых работ этого не требуется, вследствие чего картирование и сбор наиболее значимых количественных данных о растительных сообществах в реальных условиях может быть поручено даже техническим работникам, способным идентифицировать лишь основные доминирующие виды района исследований, что сильно упрощает проведение хозяйственных исследований. При этом опознаваемые описания ценотической структуры сообществ, полученные техническим персоналом, легко могут быть дополнены полноценными описаниями, сделанными специалистами-альгологами. По мнению автора, несмотря на то, что предложенные синтаксономические комбинации не основаны на флористико-фитосоциологических критериях (Вебер и др., 2005), тем не менее, принципиально они могут быть встроены в существующую иерархию фитосоциологической классификации растительности.
ТАБЛИЦА ОТКРЫВАЕТСЯ ПРИ НАЖАТИИ НА ССЫЛКУ THE TABLE CAN BE SEEN BY CLICKING ON THE LINK
Заключение В выбранном районе исследования оптимальным для описания сообществ донной морской растительности следует признать доминантно-физиономический подход. В результате конкуренции за субстрат в сообществах всегда проявляется доминирование только одного вида главного яруса, тогда как участие в сложении сообщества субдоминантов главного яруса, доминантов и субдоминантов подчиненных ярусов непостоянно. Благодаря этому, выделяемые ассоциации всегда могут быть опознаны по физиономическому облику, что удобно при проведении хозяйственных исследований. Указанное свойство позволяет описывать и классифицировать растительность при сборе данных в тяжелых условиях, при проведении съемок вдоль протяженных побережий, при сборе данных водолазами-небиологами и сотрудниками низкой квалификации, т.е. в тех случаях, когда выполнение подробных флористических сборов затруднено или невозможно. Использование выраженных численно показателей обилия и равномерных ранговых шкал позволяет получать проверяемые и сравнимые количественные данные, доступные для статистической обработки. Сведения о растительности предполагается дополнять при дальнейших исследованиях. Предложенная синтаксономическая классификация растительности может быть встроена в эколого-флористическую систему классификации. Автор сознает, что описанная методика выделения ассоциаций и описания растительности не универсальна. Очевидно, что область ее применимости будет ограничена прикладными исследованиями донной морской растительности умеренных широт. Поскольку настоящая работа имеет методический и иллюстративный характер, в качестве ближайшего ее продолжения предполагается дать подробные характеристики выделенных растительных ассоциаций вместе с обсуждением сообществ со сходным доминантным составом, описанных ранее другими авторами.
Благодарности. Автор благодарит к.б.н. И.С. Гусарову и к.б.н., доцента Д.Ф. Афанасьева за критическое обсуждение содержания настоящей статьи. Автор заявляет об отсутствии конфликта интересов, требующего раскрытия в данном сообщении.
Список литературы
Статья поступила в редакцию 24.06.2021
Об авторе Дуленин Александр Алексеевич – Alexander A. Dulenin кандадит биологических наук dulenin@mail.ru Корреспондентский адрес: 680038, Россия, г. Хабаровск, Амурский бульвар, д. 13а, ХабаровскНИРО; тел. (4212)31-54-47.
ССЫЛКА: Дуленин А.А. О выделении ассоциаций морских донных макрофитов северо-западной части Татарского пролива // Вопросы современной альгологии. 2021. № 3 (27). С. 1–17. URL: http://algology.ru/1664 DOI – https://doi.org/10.33624/2311-0147-2020-2(23)-1–17
Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно.
On the identification of marine bottom macrophyte associations in the north-western Tatar Strait Alexander A. Dulenin Khabarovsk Branch of VNIRO (Khabarovsk, Russia) Based on the analysis of algological survey materials in the northwestern Tatar Strait, a method for identifying bottom macrophyte assemblages based on the dominant-physiognomical approach was developed. An excess of abundance limits sets assemblages of each association. Each association is distinguished and named by only one, the most abundant dominant of the primary vegetation layer. To describe the associations, two abundance indicators were used: projective vegetation cover (the primary) and relative biomass (the additional). Using quantitative abundance indicators makes the сharacteristics of associations verifiable, comparable, and fit for statistical processing. The proposed methodology allows identifying vegetative assemblages unambiguously and carrying out the vegetation mapping by the physiognomical outlook of assemblages. It is especially actual during surveys in severe conditions of the far eastern and north Russian seas. Key words: Tatar strait; sublittoral; macrophyte communities identification
References
Dulenin Alexander A. Orcid ID – https://orcid.org/0000-0002-3491-6805 Khabarovsk Branch of VNIRO, Khabarovsk, Russia dulenin@mail.ru
ARTICLE LINK: Dulenin A.A. On the identification of marine bottom macrophyte associations in the north-western Tatar Strait. Voprosy sovremennoi algologii (Issues of modern algology). 2021. № 3 (27). P. 1–17. URL: http://algology.ru/1664 DOI – https://doi.org/10.33624/2311-0147-2021-3(27)-1–17
На ГЛАВНУЮ
К разделу ОБЗОРЫ, СТАТЬИ И КРАТКИЕ СООБЩЕНИЯ
|
|||
|
| ||