из Материалов Международной конференции
«Экологическая физиология водных фототрофов» I Сабининские чтения
15 ноября - 29 декабря 2012 г.
Возрастные изменения морфофункциональных характеристик представителей рода бурых водорослей Cystoseira Черного и Японского морей
Age-dependent changes of morphological and functional characteristics of the genus Cystoseira Black and Japanese seas
Камнев А.Н.1, Мессинева Е.М.1,4, Рудченко М.Н.1, Силкин В.А.2,3
Alexander N. Kamnev , Ekaterina M. Messineva, Maria N. Rudchenko,
Vladimir A. Silkin
1 - Московский государственный университет имени М.В.Ломоносова;
2 - Южное отделение Института океанологии им. П.П. Ширшова РАН, г. Геленджик-7;
3 - Институт космических исследований РАН, Москва;
4 - МАТИ-РГТУ им. К.Э.Циолковского, Москва
УДК 581.1:582.272
Проанализированы возрастные изменения морфофункциональных параметров: удельной поверхности (SS), ассимиляционного потенциала (PA), индекса автотрофности (IA) для трех видов бурых водорослей рода Cystoseira. У всех исследованных видов максимальные значения SS характерны для наиболее мелких осей высоких порядков самых молодых растений. Показано, что PA является экспоненциальной функцией порядка оси, а IA – линейной. Зависимость PA и индекса автотрофности тканей от удельной поверхности носит универсальный характер и не зависит от вида водоросли. В осенний и зимний сезоны апикальный рост замедляется и преобладающим становится рост за счет увеличения радиуса ствола.
Ключевые слова: Cystoseira; удельная поверхность(SS); ассимиляционный потенциал(PA); индекс автотрофности(IA); возрастные изменения.
Проблема морфофункциональной организации слоевищ макроводорослей достаточно давно привлекает внимание исследователей (Хайлов, Парчевский, 1983; Ковардаков и др., 1985; Силкин, Хайлов, 1988; Камнев, 1989; Хайлов и др., 1992). Главным показателем, отражающим функциональную (прежде всего продукционную составляющую) активность, является удельная поверхность слоевища (Littler, Littler, 1980; Littler et al., 1983; Карманов, Ильина, 1984, Рыгалов, 1986; Силкин, Хайлов, 1988; Камнев, 1989; Хайлов и др., 1992). Кроме того, этот показатель используется для оценки трофического статуса водоема (Миничева, 1998).
Слоевище макрофитов построено на иерархическом принципе. У макроводорослей с моноподиальным ветвлением и слоевищем цилиндрического типа на стволе формируются ветви первого порядка, на них ветви второго порядка и т.д. С повышением порядка оси удельная поверхность ветви увеличивается. Продукционные свойства слоевища и удельная поверхность имеют положительную корреляцию, но с возрастом эти параметры существенно меняются. Поэтому онтогенетический аспект морфофункциональных изменений у макрофитов привлекал и привлекает внимание исследователей (Сабинин, Щапова, 1954; Бурдин и др., 1980; Довгань, Медведева, 1982; Камнев, 1989; Камнев и др., 1986, 1988, 2006; Kamnev, Messineva, 2005).
Каждая ось таллома цистозейровых водорослей имеет свой возраст. Минимальный возраст характерен для осей наиболее высоких порядков, максимальный для оси нулевого порядка, т.е. ствола. Кроме того, в пределах каждой оси можно выделить разновозрастные участки. Очевидно, что только многолетний ствол может отражать истинное хронологическое время существования данного слоевища. Поэтому в морфофункциональной организации оси именно этого порядка необходимо искать показатели, с помощью которых можно идентифицировать как хронологический, так и физиологический возраст. Одним из таких показателей может служить изменение соотношения объемов автотрофных и гетеротрофных тканей (Kamnev, Messineva, 2005; Камнев и др., 2006). Этот показатель назван нами ассимиляционным потенциалом (PA).
В работе ставилась задача проследить возрастные изменения удельной поверхности слоевища (SS)и ассимиляционного потенциала (PA), а также установить наличие зависимости между этими параметрами. Необходимо было также выяснить влияние экологических условий на характер возрастных изменений вышеперечисленных параметров, для чего были проанализированы морфологические параметры представителей одного и того же рода из разных морей России (Черного и Японского).
Материал и методика
В качестве объектов исследования были выбраны представители рода Cystoseira C.Ag., два черноморских вида – Cystoseira barbata (Good and Wood) Ag. и Cystoseira crinita (Desf:) Bory, а также один представитель дальневосточных морей - Cystoseira crassipes (Turn.) C. Ag. (Калугина-Гутник, 1975; Петров, 1975; Перестенко, 1980). Пробы Cystoseira barbata отбирались в закрытой бухте на территории заказника Большой Утриш (примерно в 12 км от г. Анапа), а пробы Cystoseira crinita – на открытом прибойном участке у скалы Киселева (приблизительно 5 км от г. Туапсе). Отбор проб Cystoseira crassipes проводился в зоне повышенной динамики вод у мыса Проходной острова Попова залива Петра Великого Японского моря. Пробы отбирались с глубины до 1 м, что позволяло избежать светового лимитирования. Для каждого вида выборка составляла не менее 50 экземпляров.
В Черном море представители рода Cystoseira относятся к доминирующим видам и самым крупным представителям фитобентоса (Калугина-Гутник, 1973, 1975). В Японском море Cystoseira crassipes не является самым крупным и доминирующим видом, однако на некоторых участках может занимать значительные площади и является промысловым видом (Петров, 1975, Перестенко, 1980; Ковалевская, 1989).
Определение возраста слоевищ цистозейровых водорослей представляет самостоятельную задачу, и она может решаться по крайней мере двумя способами. Первый предполагает, что в год на оси нулевого порядка вырастает определенное количество ветвей первого порядка (Калугина-Гутник, 1975). Поэтому для определения возраста необходимо подсчитать общее количество ветвей на стволе и это число разделить на число, равное количеству ветвей, образующихся за один год. Второй способ основан на постоянстве среднегодового прироста ствола, и для вычисления возраста необходимо знать насколько в среднем удлиняется ствол за год (Калугина-Гутник, 1975, 1992, Ковардаков и др., 1985). Для черноморских видов C.barbata и C.crinita получены регрессионные уравнения второго порядка, связывающие возраст слоевища и его длину (Парчевский, Парчук, 1979). Для вида C. crassipes из Японского моря данные для оценки возраста отсутствуют.
Для каждого вида выделяли три возрастные группы: а) молодые – мелкие неспороносящие талломы (возраст до одного года); б) средний возраст – спороносящие 2-4-летние талломы: в) стареющие – наиболее крупные растения, преимущественно 7–9-летние. Относительный возраст отдельных элементов определяли по их удаленности от апикальной точки. Очевидно, что такое деление имеет некоторую условность. Однако существующие способы не позволяют более точно определить возраст вида.
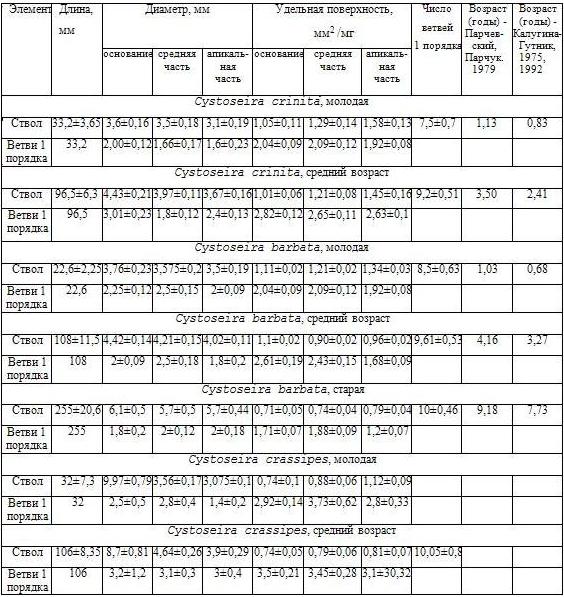
При использовании различных подходов были получены результаты, показывающие возраст водорослей, но эти данные дают возможность говорить только о приблизительной оценке возраста (Табл. 1). Молодые талломы черноморских видов имеют возраст порядка одного года. Средняя возрастная группа представлена для C. crinita 2–3-летними особями, C. barbata - 3–4-летними. Стареющая возрастная группа включает 7–9 летние талломы.
У талломов водорослей различных возрастных групп измеряли диаметры основания, средней и апикальной частей осей всех порядков, определяли их вес. Удельную площадь поверхности рассчитывали, принимая каждую часть оси за цилиндр (Табл. 1).
Для определения площадей поверхности ассимиляционного и гетеротрофного слоев свежесобранный материал фиксировали глутаровым альдегидом и заключали в аралдит (Воскобойников, Камнев, 1991). Полутонкие срезы красили гематоксилином, просматривали с помощью светового микроскопа, фотографировали и по снимкам проводили морфометрический анализ. Статистическая обработка результатов проводилась для 10%-ного уровня значимости.
Таблица 1. Морфометрические показатели осей нулевого и ветвей первого порядка у талломов различных возрастных групп трех видов цистозейр
Результаты
Теоретическая часть
Ткани осей различных порядков благодаря существующим методикам разделяются на автотрофные и гетеротрофные сегменты. Автотрофные ткани (биомасса Wа) формируют верхний слой, гетеротрофные (биомасса Wг) образуют внутреннюю часть оси. По мере увеличения диаметра оси автотрофные ткани переходят в гетеротрофные с определенной скоростью g. Динамику накопления биомассы автотрофных и гетеротрофных тканей можно описать системой дифференциальных уравнений:
dWa/dt = v•Wa – g•Wa
dWг/dt = g•Wa – m•Wг
где v - удельная скорость роста автотрофных тканей, m – удельная скорость дыхания гетеротрофных тканей.
В предельном случае, когда не происходит роста как автотрофных, так и гетеротрофных тканей, справедливо соотношение:
- Wa/ Wг = m/v; (1)
Таким образом, соотношение биомасс автотрофных и гетеротрофных тканей определяется отношением удельных скоростей дыхания гетеротрофных тканей к удельной скорости продуцирования органического вещества автотрофными тканями.
Такой показатель, как соотношение биомасс автотрофных и гетеротрофных тканей, используется для оценки физиологического состояния макрофитов и назван ассимиляционным потенциалом (PA) (Kamnev, Messineva 2005, Камнев и др., 2006). Формула (1) позволяет оценить ассимиляционный потенциал как соотношение физиологических функций – скоростей дыхания гетеротрофных тканей и продуцирования органического вещества фотосинтезирующим слоем тканей (в условиях, близких к стационарным, это равно скорости перехода автотрофных тканей в гетеротрофные).
Степень гетеротрофности или автотрофности того или иного структурного элемента таллома можно оценить с помощью другого параметра, показывающего долю автотрофных (или гетеротрофных) тканей в общей биомассе:
Ia = Wa/(Wа + Wг) = m/(m+v) = m/(m+g); (2)
Таким образом, этот показатель, который можно назвать индексом автотрофности (IA), отражает вклад дыхания гетеротрофных тканей в энергетическом балансе. Удобство этого показателя заключается в том, что выражается безразмерными единицами и заключен в интервале 0–1 (или 0–100%). При повышении скорости дыхания гетеротрофных тканей, что происходит при более высоких температурах среды, индекс автотрофности (IA) структурного элемента возрастает, повышение скорости фотосинтеза приводит к увеличению скорости перехода фотоавтотрофных тканей в гетеротрофные и, соответственно, к относительному накоплению последних.
Экспериментальные результаты
Можно ожидать, что для растений с апикальным типом роста верхушечная ткань будет более молодой, нежели ткань у основания. Это должно отразиться на морфофункциональных параметрах. В связи с тем, что очень трудно установить возраст определенного участка оси, предлагается проводить условное разделение, выделяя апикальную и среднюю части, а также основание.
Возрастные изменения в пределах ствола (оси нулевого порядка). Для всех видов водорослей характерна тенденция к увеличению такого показателя, как удельная поверхность ствола, при переходе от основания оси к апикальной части (Табл. 1). Однако достоверное увеличение этого показателя свойственно только молодым растениям. В среднем для этой возрастной группы удельная поверхность увеличивалась в полтора раза. У представителей среднего возраста достоверные различия получены только для C. crinita, для остальных видов, а также для старых талломов всех видов, тенденция увеличения не прослеживается.
Если сравнивать одинаковые части ствола у представителей разных возрастных групп одного и того же вида, то можно заметить отсутствие каких-либо изменений удельной поверхности основания. Для средней части ствола достоверное снижение удельной поверхности обнаружено только у C. barbata. Удельная поверхность апикальной части ствола снижается с возрастом у C. barbata и C. crassipes. При сравнении одинаковых частей ствола одних и тех же возрастных групп, но у разных видов, можно обнаружить, что для дальневосточной водоросли C. crassipes характерны более низкие значения удельной поверхности по сравнению с черноморскими видами.
Такой показатель, как ассимиляционный потенциал, достоверно возрастает (также примерно в полтора раза) от основания к апикальной части у молодых талломов C. crassipes и C. barbata (Табл. 2). Для других возрастных групп каких-либо тенденций не удалось обнаружить.
Таблица 2. Ассимиляционный потенциал и индекс автотрофности различных частей ствола и ветвей первого порядка у цистозейровых водорослей.
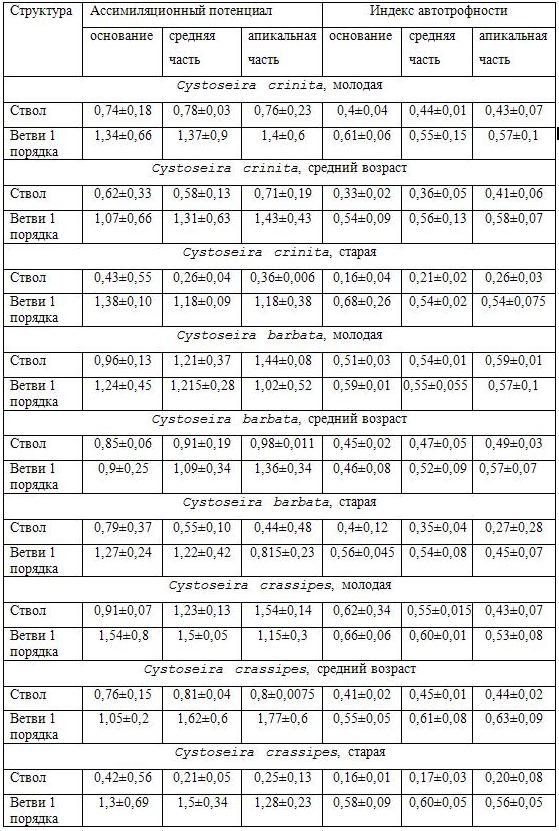
Для одинаковых частей таллома у разных возрастных групп всех исследуемых видов характерна тенденция снижения ассимиляционного потенциала по мере увеличения возраста оси. Однако для основания ствола возрастные изменения этого показателя не достоверны. В то же время в средней и апикальной частях ствола уменьшение этого показателя было существенным, в пять-шесть раз у дальневосточной C. crassipes и примерно в два-три раза у черноморских видов ассимиляционный потенциал снижается при переходе от молодых талломов к старым.
Межвидовые различия для одинаковых частей таллома у представителей одних и тех же возрастных групп в основном не прослеживаются. Исключение составляют молодые талломы C. crinita, у которых ассимиляционный потенциал средней и апикальной частей ствола ниже, чем у C. barbata и C. crassipes; у старых слоевищ C. barbata этот показатель в два раза ниже, чем у C. crinita и C. crassipes.
Для индекса автотрофности существенных изменений при переходе от старых тканей (основание) к молодым (апикальная часть) не обнаружено. Достоверное увеличение этого показателя отмечено только у старых талломов C. crinita и у молодых слоевищ C. barbata. Однако, для всех водорослей характерно снижение этого индекса по мере увеличения возраста таллома. Так, у дальневосточной водоросли C. crassipes доля автотрофных тканей основания снижается более чем в три раза при сравнении молодых и старых талломов, аналогичное сравнение для C. crinita показывает снижение более чем в два раза. У C. barbata существенных изменений доли автотрофных тканей с возрастом не обнаружено.
Межвидовые различия по такому показателю, как индекс автотрофности одних и тех же частей главной оси у особей одинакового возраста, можно охарактеризовать следующим образом. Молодые талломы C. barbata имеют более высокие значения этого показателя по всем частям ствола, нежели у других водорослей. В свою очередь значения индекса автотрофности не отличаются у стволов C. crinita и C. crassipes. Средневозрастные талломы C. crinita имеют более низкие значения этого показателя, чем аналогичные талломы C. crassipes и C. barbata. Старые талломы C. crinita и C. crassipes отличаются более низкими значениями доли автотрофных тканей.
Ветви первого порядка. Можно предположить, что ветви первого порядка, отходящие от основания, средней и апикальной частей таллома, имеют разный возраст и, как следствие, различаются по морфофункциональным показателям. При этом очевидно, что такой параметр, как удельная поверхность, будет выше у молодых ветвей первого порядка, отходящих от апикальной части, нежели у более старых ветвей, отходящих от основания оси. Однако измерения в некоторых случаях показывают противоположную тенденцию – по мере движения от основания к апикальной части в некоторых случаях наблюдается снижение этого показателя более чем на 30% (молодые талломы C. crinita, старые талломы и талломы среднего возраста C. barbata), для остальных случаев эти различия не достоверны (Табл. 1). Межвидовые различия заключаются в том, что удельная поверхность ветвей первого порядка у дальневосточной водоросли C. crassipes несколько выше, чем этот показатель у черноморских представителей. Особенно это проявляется у представителей среднего возраста.
Каких-либо закономерностей изменения ассимиляционного потенциала у ветвей первого порядка, отходящих от разных частей оси, не выявлено (Табл. 2). Также не удается установить какие-либо закономерности возрастных изменений талломов у представителей одного и того же вида.
При установлении межвидовых различий характерны более низкие значения ассимиляционного потенциала у апикальной части C. crassipes при сравнении слоевищ среднего возраста всех видов. Показатель, отражающий долю автотрофных тканей у ветвей первого порядка, существенно не зависит от места появления этой ветви на оси. Можно говорить только о некотором повышении этого показателя у старых талломов C. crinita и молодых C. barbata. Не обнаружены как межвидовые различия этого показателя для талломов одного возраста, так и различия у разновозрастных талломов одного и того же вида.
Удельная поверхность ствола и ветвей различных порядков. Формирование ветвей более высоких порядков влечет за собой увеличение удельной поверхности вплоть до уровня ветвей третьего порядка. Ветви третьего и четвертого порядков имеют практически одинаковые значения удельной поверхности, близкие 6 мм2/мг. Возрастание порядка ветвей от ствола до ветвей третьего порядка повышает удельную поверхность в среднем в 1,9 раза у дальневосточной водоросли C. crassipes, у C. crinita – в 1,4 раза, C. barbata – в 1,75 раза (Рис. 1). Эти данные приводятся для водорослей среднего возраста.
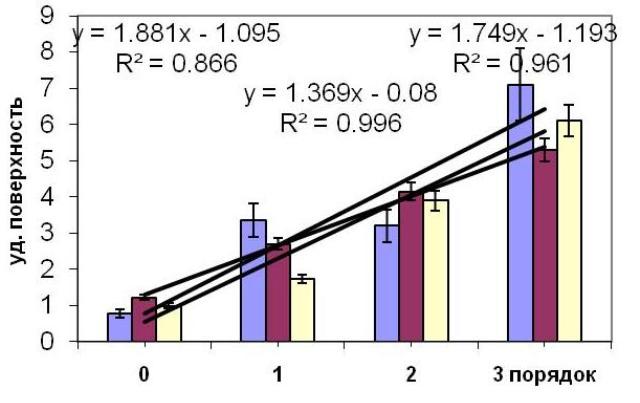

Рис. 1. Удельная поверхность ствола и ветвей первого, второго и третьего порядков цистозейровых водорослей (C. crassipes, C. crinita и C. barbata)
Такой показатель, как ассимиляционный потенциал, также существенно возрастает с увеличением порядка оси вплоть до оси третьего порядка, далее он остается в интервале 13-15. Интересной особенностью увеличения ассимиляционного потенциала с повышением порядка оси является его экспоненциальный характер (Рис. 2).
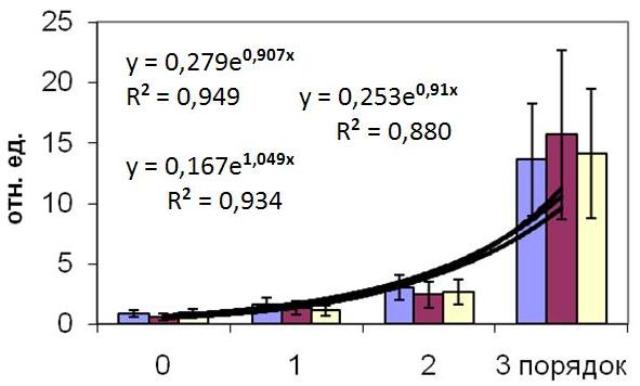
Рис. 2. Ассимиляционный потенциал ствола и ветвей первого, второго и третьего порядков цистозейровых водорослей (C. crassipes, C. crinita и C. barbata).
Y = a • exp b • X,
где X – порядок оси таллома; a и b – коэффициенты.
Попытки аппроксимировать экспериментальные данные другими зависимостями, в частности степенной и линейной, показали более низкие значения коэффициента детерминации.
Индекс автотрофности оси также возрастает с увеличением порядка оси вплоть до оси третьего порядка, далее остается на том же уровне, равном 0,93. Причем эта зависимость носит линейный характер, в среднем доля автотрофных тканей повышается у талломов среднего возраста в 1,55 – 1,83 раза при переходе к оси более высокого уровня (Рис. 3).
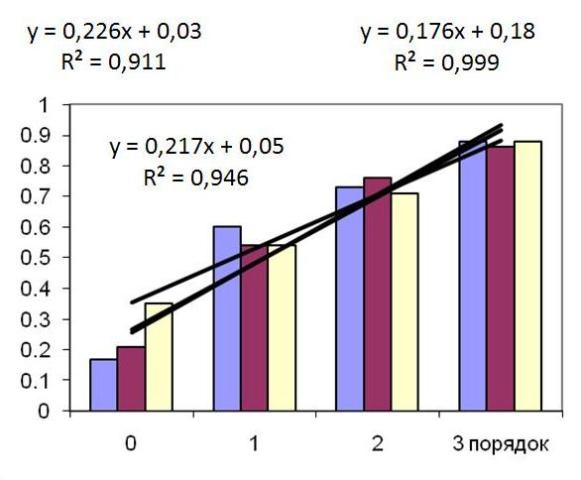
Рис. 3. Индекс автотрофности ствола и ветвей первого, второго и третьего порядков цистозейровых водорослей (C. crassipes, C. crinita и C. barbata)
Как влияет возраст таллома на закономерности изменения ассимиляционного потенциала и на индекс автотрофности при увеличении порядка оси? Для этого следует рассмотреть изменение коэффициентов экспоненциального уравнения, описывающего изменение ассимиляционного потенциала с возрастом таллома (Табл. 3).
Таблица 3. Коэффициенты экспоненциального уравнения, описывающего изменения ассимиляционного потенциала с увеличением порядка оси для талломов различного возраста.
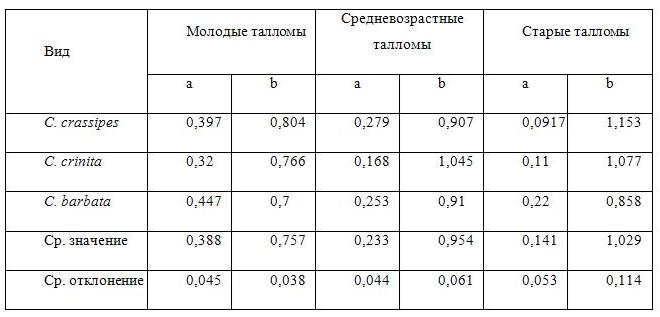
Общей тенденцией является уменьшение в 2–4 раза параметра a, отражающего значение ассимиляционного потенциала ствола таллома и повышение значений коэффициента b на 20-30% при переходе от молодых талломов к старым. Это указывает на то, что с увеличением возраста таллома происходит существенное снижение ассимиляционного потенциала ствола и ветвей низших порядков.
Интересные особенности проявляются у параметра, отражающего индекс автотрофности оси у таллома разного возраста, изменение которого лучше описывается линейной зависимостью (Табл. 4):
Y = a + bX,
где X – порядок оси таллома, a и b – коэффициенты.
Таблица 4. Коэффициенты линейного уравнения, описывающего изменения доли автотрофных тканей с увеличением порядка оси для талломов различного возраста.
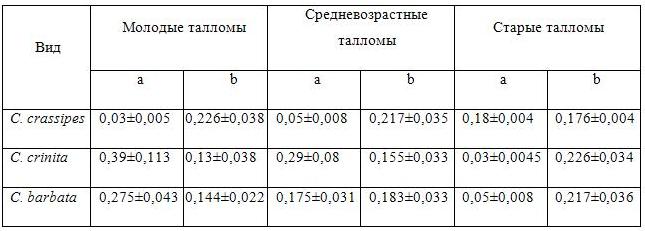
У дальневосточной водоросли C. crassipes коэффициент a увеличивается с возрастом таллома, а коэффициент b имеет тенденцию к снижению. У черноморских видов наблюдается обратная тенденция (снижение коэффициента a и увеличение коэффициента b).
Обсуждение результатов
Рост биомассы макрофитов с моноподиальным слоевищем цилиндрического типа определяется тремя степенями свободы:
1) рост длины оси путем формирования новых клеток на апикальной части,
2) рост в толщину и, как следствие, повышение радиуса цилиндра, за счет латерального деления и роста клеток
3) образование на данной оси ветвей более высоких порядков.
Первые два типа роста характеризуют экстенсивный путь образования биомассы, третий – интенсивный. Очевидно, преобладание того или иного типа роста биомассы зависит от условий окружающей среды. Удельная поверхность таллома возрастает только при реализации интенсивной стратегии роста. При росте оси в длину удельная поверхность остается на прежнем уровне, повышение толщины цилиндра приводит к уменьшению данного морфологического показателя. Проявившиеся достоверные различия в удельной поверхности между основанием и апикальной частью у молодых растений говорят о том, что в первый год у растений преобладает рост длины оси. В осенний и зимний сезоны апикальный рост замедляется и преобладающим становится рост биомассы за счет увеличения радиуса ствола. В последующие годы эти два пути роста биомассы ствола становятся сбалансированными и, поэтому, не удается обнаружить четких различий удельной поверхности основания и апикальной части. В первый год также происходит окончательное формирование толщины ствола, в дальнейшем этот показатель изменяется незначительно, что не позволяет проследить изменений удельной поверхности одинаковых частей ствола у представителей разных возрастных групп. Необходимо заметить, что аналогичный характер изменений толщины ствола характерен и для других макрофитов, в частности, красной черноморской водоросли грацилярии (Силкин, Миронова, 2005).
Обращает на себя внимание тот факт межвидовых различий, что для дальневосточной водоросли C. crassipes характерно более низкое значение удельной поверхности, чем для черноморских видов C.crinita и C. barbata. Предельное значение удельной поверхности (S/W) определяется соотношением двух параметров (Силкин и др., 1992):
S / W = q /Io;
q – поток энергии, идущий на поддержание структуры;
Io – поток световой энергии, падающий на поверхность.
Поскольку водоросли растут в условиях одной и той же географической широты, поток световой энергии в обоих случаях будет приблизительно равным. Однако поток энергии, идущий на поддержание, зависит от температуры, существенно увеличиваясь при ее повышении. В естественных местах обитания в летний период дальневосточная водоросль C. crassipes растет при температуре примерно на 6–10о С ниже, чем черноморские виды C. crinita и C. barbata. Возможно, что это позволяет дальневосточному виду достигать большей толщины ствола и, соответственно, меньших значений удельной поверхности.
Данное соотношение позволяет объяснить, почему ветви первого порядка, отходящие от апикальной части, в некоторых случаях имеют большую толщину и, как следствие, меньшую удельную поверхность. Это связано с тем, что ветви у апикальной части получают больше световой энергии из-за меньшего затенения.
Ассимиляционный потенциал различных частей ствола молодых растений достоверно отличается, что указывает на необходимость времени для перехода автотрофных тканей в гетеротрофные, и чем старше часть ствола, тем более низким становится значение этого функционального показателя. Возраст таллома существенно определяет величину ассимиляционного потенциала. При переходе от молодых талломов к старым отмечено более существенное снижение данного показателя. Это указывает на то, что превращение автотрофных тканей в гетеротрофные происходит в течение длительного времени, сравнимого с возрастом слоевища.
Более низкие значения ассимиляционного потенциала и индекса автотрофности у дальневосточной водоросли C. crassipes по сравнению с черноморскими видами можно также объяснить в соответствии с формулами 1 и 2 более низкими величинами удельных скоростей дыхания – m.
Температурные условия среды, по-видимому, определяют разнонаправленность возрастных изменений коэффициентов линейного уравнения для дальневосточного и черноморских видов (Табл. 4).
Очевидно, что ассимиляционный потенциал связан с удельной поверхностью, поскольку эти параметры зависят от радиуса оси. Действительно, обнаружена линейная корреляция между логарифмом ассимиляционного потенциала и удельной поверхностью (Рис. 4).
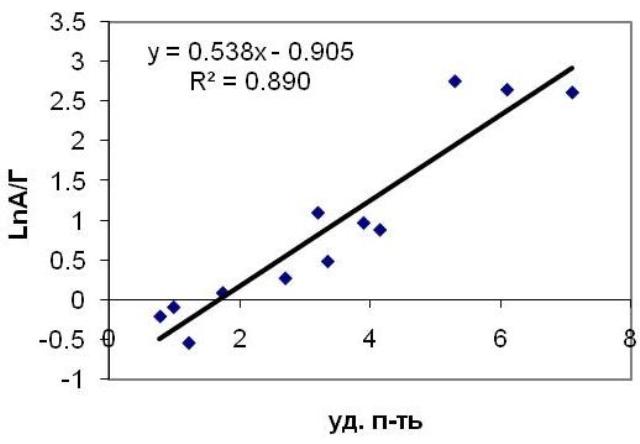
Рис. 4. Соотношение между ассимиляционным потенциалом и удельной поверхностью для исследуемых видов водорослей.
Предположение о связи между параметром, отражающим долю автотрофных тканей, и удельной поверхностью также нашло подтверждение (Рис. 5).
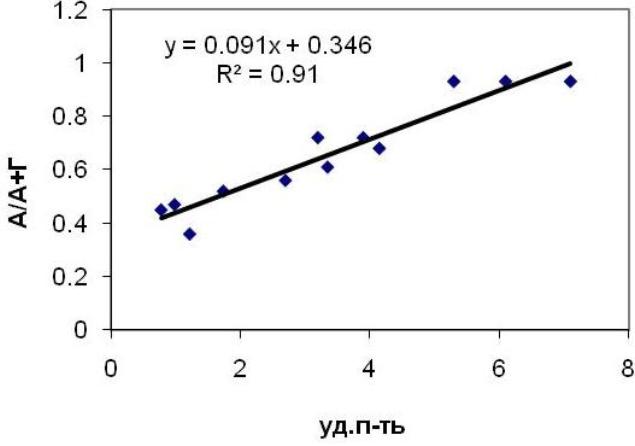
Рис. 5. Соотношение между индексом автотрофности тканей и удельной поверхностью для исследуемых видов водорослей.
Следует заметить, что в обоих случаях объединены экспериментальные данные по трем видам водорослей. Это указывает на то, что зависимость ассимиляционного потенциала и доли автотрофных тканей от удельной поверхности носит универсальный характер и не зависит от вида водоросли. Отсюда следует, что по удельной поверхности можно с определенной долей вероятности прогнозировать ассимиляционный потенциал (PA) и индекс автотрофности (IA) как отдельного структурного элемента, так и таллома в целом.
В связи с тем, что водоросли с моноподиальным строением состоят из структурных элементов не только разного хронологического, но и физиологического возраста, то такой организм представляет собой совокупность органов различного физиологического возраста. Именно это и обеспечивает функционирование таллома в течение длительного времени с достаточно высоким продукционным потенциалом.
Список литературы
1. Бурдин К.С., Гусев М.В., Крупина М.В., Савельев И.Б. Изучение возможности использования макроводорослей в качестве организма-монитора загрязнения Чёрного моря тяжёлыми металлами // Вестник МГУ. 1980. Т.16, №3. С.3-11.
2. Воскобойников Г.В., Камнев А.Н. Морфо-функциональные изменения хлоропластов в онтогенезе водорослей. Спб.: Наука, 1992. 96 с.
3. Довгань И.В., Медведева Е.И. Гистохимическое изучение бурой водоросли Cystoseira barbata разного возраста // Гидробиол. ж. 1982. Т.18, №5. С.95-99.
4. Калугина-Гутник А.А. Некоторые особенности развития черноморской цистозиры (Cystoseira barbata и Cystoseira crinita) // Бот.ж. 1973. Т.58, №1. С.20-30.
5. Калугина-Гутник А.А. Сезонная динамика роста Cystoseira barbata (Good. et Wood.) Ag. (Phaeophyta) в Чёрном море // Альгология. 1992. Т. 2. №1. С.32-40.
6. Калугина-Гутник А.А. Фитобентос Чёрного моря. Киев: Наук. думка, 1975. 248 с.
7. Камнев А.Н. Структура и функции бурых водорослей. М.: МГУ, 1989. 199 с.
8. Камнев А.Н., Золотухина Е.Ю., Бурдин К.С. Возрастные изменения некоторых физиологических характеристик бурой водоросли Sargassum pallidum // Физиол. раст. 1986. Т.33. С.930-934.
9. Камнев А.Н., Бурдин К.С., Камнева М.А., Родова Н.А. Анатомическое строение разновозрастных слоевищ бурой водоросли Sargassum pallidum (Turn.) C. Ag. // Изв. АН СССР. 1988. № 11. С.301 –305.
10. Камнев А.Н., Мессинева Е.М.,Силкин В.А. Использование новых коэффициентов при исследовании морфофункциональных характеристик водорослей // Материалы международной конференции посвященной 75-летию Биологического факультета МГУ им.М.В.Ломоносова. - М., Макс Пресс, 2006. C.92-97.
11. Карманов И.В., Ильина Н.А. Взаимосвязь между пространственной структурой и продукцией на разных уровнях организации (особь, ярус, сообщество) // Ботан. журн. 1984. 69, № 12. С.1593–1602.
12. Ковалевская Р.А. Морфология, размножение, экология Cystoseira crassipes (Turn.) C. Ag.: дисс. ... канд. биол. наук. - Владивосток, 1989. - 181с.
13. Ковардаков С.А., Празукин А.В., Фирсов Ю. К., Попов А.Е. Комплексная адаптация цистозиры к градиентным условиям: научные и прикладные проблемы. Киев: Наук. думка, 1985. 216 с.
14. Парчевский В.П., Парчук Г.В. Анализ морфологических признаков черноморских цистозир в онтогенетическом ряду в природных условиях // 3 Всесоюзное совещание по морской альгологии – макрофитобентосу: тез. докл. Киев: Наукова думка, 1979. С.96-98.
15. Перестенко Л.П. Водоросли залива Петра Великого. Л.: Наука, 1980. 232 с.
16. Петров Ю.Е. Ламинариевые и фукусовые водоросли морей СССР (морфология, экология, филогения, систематика): дис. … докт. биол. наук. - Л.: БИН АН СССР, 1975. 287 с.
17. Рыгалов В.Е. Теоретико-экспериментальный анализ роста морских макрофитов (на примере Ahnfeltia tobuchiensis): Автореф. канд. диссер. канд. биол. наук. - Красноярск: Институт биофизики, 1986. - 23 с.
18. Сабинин Д.А., Щапова Т.Ф. Темп роста, возраст и продукция Cystoseira barbata в Чёрном море // Тр. Ин-та океанологии. 1954. Т.8. С.119-146.
19. Силкин В.А., Хайлов К.М. Биоэкологические механизмы управления в аквакультуре. Л.: Наука, 1988. 230 с.
20. Силкин В.А., Миронова, Н.В. Динамика продукционных и морфологических показателей таллома грацилярии в культуре // Альгология. 2005. 15, №1. С.14-27.
21. Силкин В.А., Золотухина Е. Ю., Бурдин К.С. Биотехнология морских водорослей. М.: Изд-во МГУ. 150 с.
22. Хайлов К.М., Парчевский В.П. Иерархическая регуляция структуры и функции морских растений. Киев.: Наук. думка, 1983. 254 с.
23. Хайлов К.М., Празукин А.В., Ковардаков С.А., Рыгалов В.Е. Функциональная морфология морских многоклеточных водорослей. Киев: Наук. думка, 1992. 280 с.
24. Kamnev A.N., Messineva E.M. New simple coefficients for macroalgal functional assessments // Phycologia. V.44. № 4. 2005. Р.51.
25. Littler M.M. and Littler D.S. Morphological form and photosynthetic performances of marine macroalgae: tests of a functional-form hypotheses // Bot. Mar. 1980. 22. P.161-163.
26. Littler M.M., Littler D.S., Taylor, P.R. Evolutionary strategies in tropical barrier reef system: functional- form groups of marine macroalgae // J. Phycol. 1983. V. 19. P.229–237.
опубликовано - декабрь 2012 г.
Age-dependent changes of morphological and functional characteristics of the genus Cystoseira Black and Japanese seas
Alexander N. Kamnev , Ekaterina M. Messineva, Maria N. Rudchenko, Vladimir A. Silkin
Three brown algae species from genus Cystoseira (C.barbata, C.crinita, C.crassipes) were used as model for studying of some morphological and functional characteristics age changing. The value of specific surface - SS (mm2/mg) is decreasing with age. Some differences between species are also found. The assimilative potential - PA (ratio of assimilative / heterotrophic amount of tissue) is increasing a little, but the level of this factor is nearly equal in all studied species. Shown that assimilative potential (PA) is an exponential function of axis level, avtotrofnosti index (IA) is linear. Dependence of the potential of assimilative (PA) and autotrophic tissues of specific surface (SS) is universal and does not depend on the type of algae. It follows that the specific surface (SS) is possible with a certain probability to predict assimilative potential (PA), and the avtotrofnosti index (IA) as a structuring element and tallus in general.
Key words: Cystoseira; specific surface(SS); assimilative potential (PA); avtotrofnosti index(IA); age changing.
Об авторах
Камнев Александр Николаевич - Kamnev Alexander N.
доктор биологических наук
ведущий научный сотрудник ФГБОУ ВПО «Московский государственный университет им.М.В.Ломоносова», Москва, Россия (Lomonosov Moscow State University, Moscow, Russia), Биологический факультет, кафедра физиологии растений
dr.kamnev@mail.ru
Мессинева Екатерина Михайловна - Messeneva Ekaterina M.
кандидат биологических наук
ФГБОУ ВПО «Московский государственный университет им.М.В.Ломоносова», Москва, Россия (Lomonosov Moscow State University, Moscow, Russia); МАТИ-РГТУ им. К.Э.Циолковского, Москва, Россия (MATI - Russian State Technological University, Moscow, Russia).
musculus@mail.ru
Рудченко Мария Николаевна - Rudchenko Maria N.
аспирант
ФГБОУ ВПО «Московский государственный университет им.М.В.Ломоносова», Москва, Россия (Lomonosov Moscow State University, Moscow, Russia), Биологический факультет, кафедра микологии и альгологии.
mashencija@list.ru
Силкин Владимир Арсентьевич - Silkin Vladimir A.
доктор биологических наук
главный научный сотрудник Южного отделения ФГБУН «Институт океанологии им.П.П.Ширшова РАН», Геленджик, Россия (Southern branch of the P.P. Shirshov Institute of Oceanology RAS, Gelendzhik, Russia); Институт космических исследований РАН, Москва (Space Research Institute RAS, Moscow, Russia)
vsilkin@mail.ru
Корреспондентский адрес: Россия, 119991, Москва, Ленинские горы, Московский Государственный Университет им.М.В.Ломоносова, д. 1, стр. 12, Биологический ф-т, каф. физиологии растений; тел.: (495) 939-21-18.
ССЫЛКА НА СТАТЬЮ:
Камнев А.Н., Мессинева Е.М., Рудченко М.Н., Силкин В.А. Возрастные изменения морфофункциональных характеристик представителей рода бурых водорослей Cystoseira Черного и Японского морей // Вопросы современной альгологии. 2012. № 2 (2). URL: http://algology.ru/148
К другим статьям Международной конференции «Экологическая физиология водных фототрофов». I Сабининские чтения
Карта сайта

|
