из Материалов Международной конференции
«Экологическая физиология водных фототрофов» I Сабининские чтения
15 ноября - 29 декабря 2012 г.
Макрофитобентос как индикатор динамики подводных ландшафтов береговой зоны моря
Macrophytobenthos as an indicator of bottom marine seascapes in coastal zone
Папунов Д.В.
Dmitry V. Papunov
Государственный океанографический институт им. Н.Н. Зубова
УДК 574.587
В работе рассмотрены основные механизмы динамики подводных ландшафтов береговой зоны. В качестве индикатора состояний подводных ландшафтов предложено использование биомассы и численности видов макрофитобентоса. В качестве примера предложены тестовые полигоны исследований на Черном и Белом морях. Выявлены основные закономерности сезонной динамики биомассы макрофитов в зависимости от глубины.
Ключевые слова: подводные ландшафты; индикатор ландшафтной динамики; фитобентос береговой зоны моря; Черное море; Белое море; динамика подводных ландшафтов.
Интерес к изучению береговой зоны моря объясняется расширением хозяйственного использования ее природных ресурсов, что требует детального анализа структуры подводных ландшафтов и их динамики. Основы исследований в этой области географии заложены в работах К.К. Маркова, Е.Ф. Гурьяновой, В.В. Шаркова, К.М. Петрова.
Преобладающая часть современных исследований подводных ландшафтов, или донных природных комплексов (ДПК) ориентирована на изучение их пространственной структуры, оставляя в стороне вопросы динамики. Исследование явлений и процессов без учета их временной изменчивости является недостаточным, поскольку отсутствие данных о динамике исследуемых ДПК в значительной степени затрудняет возможности прогноза последствий реализуемых хозяйственных проектов. Таким образом, для оценки природно-ресурсного потенциала и выработки стратегии устойчивого использования береговой зоны моря и для разработки регламентов ее охраны представляется целесообразным использование ландшафтных методов.
В настоящее время многие авторы отмечают актуальность изучения динамики подводных ландшафтов, предпринимаются попытки осмысления и изучения динамики подводных ландшафтов. Однако, ввиду ограниченного опыта подобных исследований, методологических основ и накопленного массива данных, изучение динамики ландшафтов сводится к изучению динамики их компонентов.
Подводный ландшафт, как и ландшафт сухопутный есть сочетание абиотических и биотических составляющих. К первым относятся рельеф, донные осадки, гидрологический и гидродинамический режимы, содержание микроэлементов в толще воды и осадках. К биотическим - донное население и растительность, содержание биогенов в толще воды.
Среди факторов ландшафтной динамики выделены активные и пассивные факторы. Активные включают процессы, преобразующие литогенную основу подводного ландшафта (гидродинамика, ледовые условия). Пассивные факторы, помимо характера донных отложений и рельефа, включают различные гидрохимические и гидрофизические характеристики водной массы. Температура, соленость, прозрачность, состав и количество взвешенных и растворенных веществ определяют биологическую составляющую ландшафта, т.е. возможность существования в данном районе тех или иных видов подводных организмов.
Одним из основных индикаторов состояния донного природного комплекса является бентосная растительность. Динамика состояний фитобентоса может быть связана с изменениями, вызванными внешними факторами, такими как: изменения подстилающего материала (грунта), гидродинамической нагрузки, уровня освещенности, химических (соленость, содержание растворенных веществ) и физических (температура, содержание взвешенных частиц) характеристик вод. Как правило, эти изменения проходят в многолетний период. Кроме этого, имеют место цикличные изменения состояний фитоценозов, вызванные сменой времен года.
Анализ литературы и опыт полевых исследований показали, что для большинства средне- и высокоширотных морей северного полушария, существуют сезонные различия в состояниях ассоциаций макрофитов. Сезонные изменения состояния фитобентоса обусловлены внутригодовым ходом температуры, освещенности, волновой нагрузкой и другими параметрами окружающей водной массы. Эти различия отражаются в циклах жизни многолетних растений: периодах размножения, наибольшей биомассы, отмирания и т.п. Не углубляясь в экологию водорослей-макрофитов, отметим, что для всех водорослей и ассоциаций эти периоды различны по времени наступления и продолжительности. Кроме того, для некоторых морей имеют место сезонные различия в антропогенной нагрузке на экосистемы. Так, например, в весенне-летний период Черное море подвергается наибольшему антропогенному воздействию, которое связано с началом курортного сезона.
Многолетние состояния водорослевых ассоциаций и их сукцессии могут быть связаны как с природными (изменение климата, изменение уровня моря и др.), так и с антропогенными факторами: качеством вод, добычей полезных ископаемых, добычей водорослей.
Важным вопросом при разработке методики изучения динамики ДПК является вопрос об индикаторах динамики ДПК береговой зоны. В качестве индикаторов динамики ДПК могут выступать как изменения отдельных свойств одного или нескольких компонентов подводного ландшафта (консолидированность и гранулометрический состав донных отложений, видовой состав бентосных организмов, интенсивность потока энергии между компонентами и проч.), так и эмерджентные свойства всего природного комплекса: структура, устойчивость, продуктивность, пригодность для того или иного вида хозяйственной деятельности и др. В связи с тем, что эмерджентные свойства подводных ландшафтов на сегодняшний день изучены слабо, в качестве индикаторов ландшафтной динамики могут быть приняты различные компоненты подводного ландшафта.
Как отмечалось, одним из основных внутренних природных факторов ландшафтной динамики в береговой зоне моря является изменчивость гидроклиматического режима вод. Следствием этих явлений выступает изменчивость литогенной основы (рельефа и отложений) и биотического компонента ландшафта. Таким образом, можно говорить о двух типах индикаторов динамики подводных ландшафтов в береговой зоне: биологических и геоморфологических. При этом необходимо отметить, что действие биологических индикаторов проявляется в основном при смене внутрисезонных, сезонных, годовых и многолетних состояний донного природного комплекса, а действие геоморфологических – при смене годовых и многолетних состояний. Как правило, при этом геоморфологический индикатор выступает более инертным по отношению к биотическому индикатору. Естественно, это утверждение не относится к тем участкам дна береговой зоны моря, где массовые поселения бентоса отсутствуют. К таковым относятся, например, отмелые аккумулятивные участки сложенные песчаными отложениями, характеризующиеся интенсивным волновым воздействием.
Биотические индикаторы состояний ДПК можно подразделить на фитоиндикаторы и зооиндикаторы. К ним относятся бентосные животные (зооиндикаторы) и растения (фитоиндикаторы), которые обладают легко обнаружимыми особенностями, проявляющимися при смене окружающей среды: температуры, освещенности, химического состава, гидродинамики вод; состава, консолидации и других свойств донных отложений. Индикационными способностями могут обладать как сообщества бентоса, так и отдельные виды.
Для каждого определенного изменения окружающей среды существуют отдельные индикаторные виды и/или сообщества бентосных организмов. Важно отметить, что при определении биологических индикаторов необходимо руководствоваться тем, что индикаторные свойства индикаторов должны быть физиономическими, то есть визуально определяемыми, так как зачастую многие изменения в бентосных организмах и структуре их сообществ невозможно определить без специальных мероприятий (напр. химических анализов) и приборов (микроскопов и проч.).
Донные сообщества водорослей четко реагируют на изменения косной среды: температуры, освещенности, солености, гидродинамической нагрузки, загрязнения и др. Как отмечает К.М. Петров (Комплексное физико-географическое изучение морских мелкводий, 1973) (Петров, 1973): «Подводная растительность является той ячейкой геосферы, в которой ассимилируется основная часть солнечной энергии, продуцируется органическое вещество и осуществляется обмен веществом и энергией с другими геосистемами. Подводная растительность, чутко реагируя на комплекс экологических факторов, определяющих специфику местообитания, является хорошим индикатором своеобразия морфологических комплексов ландшафтов морских мелководий».
Основные изменения бентосных сообществ водорослей выражаются во флористическом составе ассоциаций, интенсивности роста, сроках вегетации, биомассе, а также первичной продукции.
В качестве тестовых полигонов изучения динамики подводных ландшафтов был использован участок береговой зоны Черного моря: мыс Малый Утриш (п-ов Абрау) и Белого моря - залив Нильмогуба (Кандалакшский залив). При их выборе основным требованием являлось различие в природных условиях: находиться в различных географических зонах, быть сопредельным с различными типами берегов, находиться в условиях моря с выраженными и невыраженными приливно-отливными явлениями.
Изучение ландшафтной структуры и динамики выбранных участков проводилось при использовании водолазного оборудования. Ежегодно с 2005 по 2008 г. в летний и зимний периоды проводилась подводная ландшафтная съемка на выбранных участках методом ландшафтного профилирования. Для фиксирования сезонных состояний ДПК нами проводились измерения биомассы водорослей макрофитов. Для этого производился сбор водорослей на площади 1 м2, в характерных ДПК. Взвешивание водорослей с целью определения биомассы осуществлялось при помощи хозяйственных весов. Вопреки методике, принятой в альгологии, мы пренебрегли использованием рамки, в связи с тем, что при мозаичном распространении водорослей, измерения при помощи рамки даёт большие погрешности.
На основе полученных карт подводных ландшафтов полигонов построены карты-схемы сообществ донных фитоценозов (рис. 1 и 2).
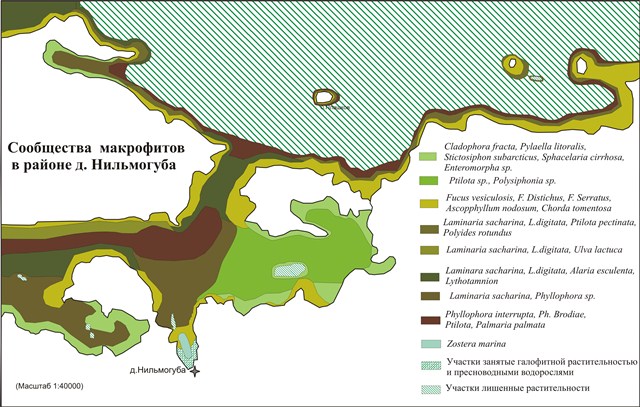

Рис.1. Сообщества макрофитов в районе д.Нильмогуба (Белое море)
(ПП - проективное покрытие)
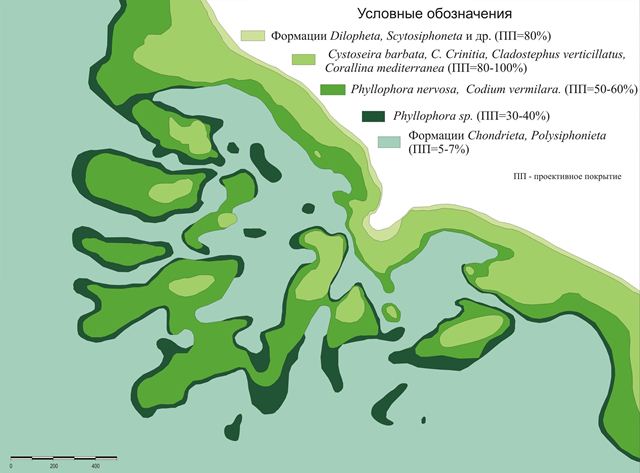
Рис.2. Сообщества макрофитов в районе м. Малый Утриш (Черное море)
Во время исследований на черноморском полигоне исследований выявлено, что в летний сезон в бентосных фитоценозах наблюдается наибольшая биомасса и высокое видовое разнообразие. Это достигается за счет появления в ассоциациях многолетних водорослей сопутствующих сезонных однолетних видов, а также интенсивного роста большинства многолетних макрофитов.
В мелководных ДПК, приуроченных к прибойной зоне (от 0,5 до 2-2,5 м), в летний период появляется большое количество сезонных водорослей: Dilophus fasciola, Enteromorpha compressa, Ceramium cilitum, Corallina officinalis, Cladostephus verticillatus и др. Биомасса макрофитов летом составляет около 500-600 г/м2, а количество видов достигает 40.
Глубже (от 2 до 9 м), в ассоциации Cystoseira crinitia+C.barbata – Cladostephus verticillatus – Corallina meditarrenea, в летний период наблюдаются наибольшие значения биомассы. Видовой состав наиболее богат в конце весны и особенно летом, что связано с массовым появлением сезонных летних форм (Polysiphonia sp., Ceramium sp., Enteromorpha sp. и др.). Биомасса обеих видов цистозиры на различных участках составляет 75-96% от общей биомассы макрофитов ассоциации. Биомасса сопутствующих видов приходится на весенне-летний период, когда эпифитные синузии и ряд сопутствующих форм достигают максимального развития. Общая биомасса макрофитов в этом поясе (в районе исследований) летом может достигать 5 кг/м2, а количество видов – около 50.
Кроме появления или исчезновения сезонных или однолетних видов, сезонные изменения условий произрастания выражены в скорости роста многолетних водорослей, росте отдельных органов, например у цистозиры летом наблюдается рост генеративных ветвей. В сезонной динамике биомассы цистозиры наблюдается два максимума: весенний (апрель-июнь) и осенний (сентябрь-ноябрь), что связано с интенсивным ростом в эти периоды и массовым развитием репродуктивных органов (Калугина-Гутник, 1975). Наименьшая биомасса этой водоросли приходится на лето и зиму, когда рост слоевищ замедляется, а большинство ветвей, выросших за весенний и осенний период, опадает.Для более глубоководных сообществ Phillophora sрp. и Codium vermilara конец весны и начало лета является периодом наибольшей биомассы (около 0,6 кг/м2) и наибольшего видового разнообразия (30 видов) (Калугина-Гутник, 1975).
Для характеристики изменчивости состояний бентосных фитоценозов, как основных индикаторов состояний ДПК, приведены процентные соотношения зимней и летней биомассы, а также видового разнообразия летом и зимой (рис. 3). При их анализе видно, что основные изменения биомассы в бентосных фитоценозах приурочены к прибойной зоне, и с увеличением глубины они уменьшаются. Количественные различия в видовом составе макрофитов наименьшие на малых глубинах, это связано с развитием как летних так и зимних сезонных форм водорослей, что приводит к постоянному общему количеству видов.
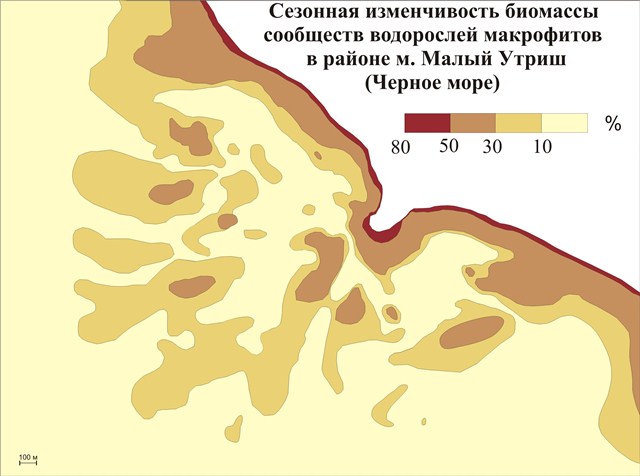
Рис.3. Сезонная изменчивость биомассы сообществ водорослей макрофитов в районе м. Малый Утриш (Черное море)
На Беломорском полигоне исследований для водорослей-макрофитов периодом активного роста является весна-лето, время с наибольшим видовым разнообразием и биомассой, а периодом покоя – осень-зима, для которого характерно меньшее видовое разнообразие и соответственно меньшая биомасса. Различия в летних и зимних значениях биомассы и количества видов неодинаковы по глубине и наибольшие из них приурочены к поясу фукоидов и нитчаток (от 0 до до 2 м) и ламинариево-аляриевому поясу (от 2 до 6-8 м).
Для характеристики сезонных изменений построена карта процентного соотношения значений летней и зимней биомассы водорослей полигона (рис. 4). Ее анализ показывает, что максимальные сезонные изменения биомассы (до 100%) наблюдаются в интервале глубин от 0 до 8 м, наименьшие – на глубинах более 15 м. С увеличением глубины сезонные изменения постепенно сглаживаются и к глубинам 30-40 м практически исчезают. Это связано с тем, что на данных глубинах освещенность во время всего года практически отсутствует, а амплитуды температур почти не выражены. На мелководье наиболее слабым сезонным изменениям подвержены ДПК узких проливов и морских порогов, в тех местах, где интенсивные приливные течения не дают сформироваться ледовому покрову.
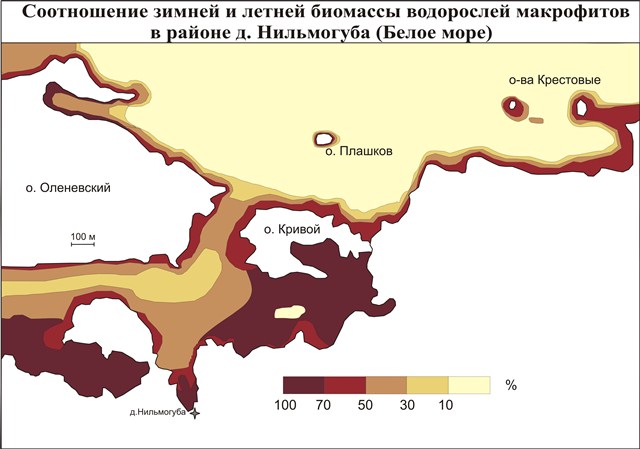
Рис.4. Соотношение зимней и летней биомассы водорослей макрофитов в районе д.Нильмогуба (Белое море)
Анализируя полученные данные, можно говорить о том, что максимальные изменения состояний ДПК на Белом море присущи мелководным участкам. Наибольшим изменениям подвержены мелководные участки, на которых образуется устойчивый ледяной покров. Отсюда можно утверждать, что наиболее значимым фактором сезонной динамики беломорских ДПК является образование устойчивого ледового покрова, следствием чего является прекращение волнового воздействия на дно и значительное снижение освещенности.
Различные физико-географические условия полигонов и ландшафтная структура определяют особенности ландшафтной динамики. Так, основными факторами сезонной динамики на черноморском полигоне является изменчивость интенсивности ветрового волнения и температуры, а на беломорском полигоне – температуры, освещенности и ледовой абразии. Одной из общих черт сезонной динамики обоих полигонов является зависимость ее интенсивности от глубины: с глубиной различия в летних и зимних состояниях уменьшаются. Сезонная динамика ДПК беломорского полигона затухает на 5 м ближе к поверхности, по сравнению с Черным морем, что связано с развитием зимой на Белом море ледового покрова и низкой инсоляцией.
Кроме сезонной динамики, донные макрофиты могут служить индикатором многолетней динамики подводных ландшафтов. Так, в качестве индикатора многолетней динамики ДПК черноморского полигона исследований выбрана водоросль цистозира (Cystoseira barbata, C. crinitia), которая представляет наиболее распространенную водоросль Черного моря и является важным промысловым видом.
Опираясь на литературные данные и собственные наблюдения, мы можем выделить некоторые изменения в ландшафтной структуре ДПК черноморского полигона исследований. В основном эти изменения проявляются в горизонтальной структуре растительного покрова.
Как отмечает Е.И. Блинова (2007), за последние 20-40 лет экосистема Черного моря претерпела существенные изменения под влиянием антропогенного воздействия: зарегулирования стока рек, увеличения сброса загрязненных вод промышленными и сельскохозяйственными предприятиями, бытовых стоков и других. В результате антропогенного пресса произошли значительные изменения гидрохомического режима моря, что вызвало перестройку донных сообществ: сокращение площадей фитобентоса в основном за счет поднятия на меньшие глубины нижней его границы, изменение видового состава, снижение биомассы, прежде всего многолетних видов, и значительное уменьшение их запасов (Блинова, 2007).
Так, по данным (Промысловые водоросли СССР, 1971) нижняя граница распространения массовых поселений цистозиры (Cystosiera barbata) в районе наших исследований 35-40 лет назад проходила на глубине около 32 м. На основе этих данных и выявленной ландшафтной структуре построена карта распространения цистозировых сообществ на 1971 г. (рис. 5).
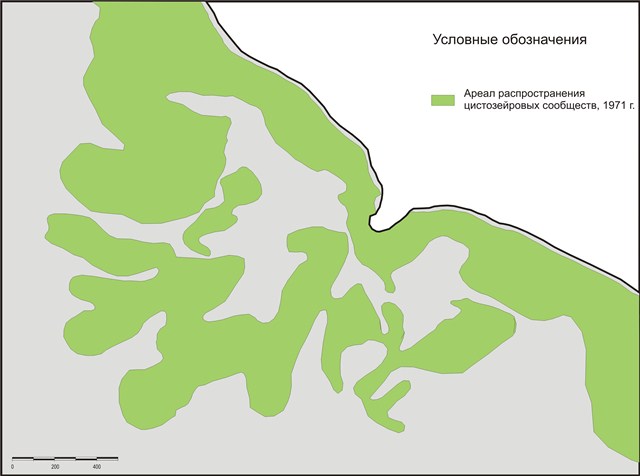
Рис.5. Карта распространения цистозировых сообществ на 1971 г. (Черное море, район м. Малый Утриш)
В результате проведенных исследований, выявлено: на сегодняшний день нижняя граница распределения массовых поселений цистозиры на м. Малый Утриш расположена на глубине 8-9 м (рис. 6), что подтверждает многочисленные данные о сокращении ареала ее распространения в результате антропогенного пресса (в частности общей эвтрофикации вод Черного моря), а также некоторых природных процессов – увеличения численности гребневика мнемиопсиса (Mnemiopsis leidyi), который, питаясь зоопланктоном, нарушил трофическую цепь в экосистеме и дал толчок к неконтролируемому размножению фитопланктона, массовое развитие которого, в свою очередь, стало причиной снижения прозрачности вод и, соответственно, сужения зоны фитали.
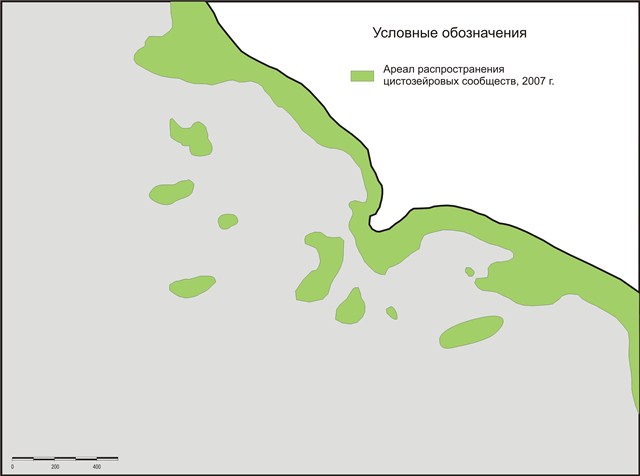
Рис.6. Карта распространения цистозировых сообществ на 2007 г. (Черное море, район м. Малый Утриш)
В заключение необходимо отметить, что донные сообщества макрофитов являются не единственным возможным индикатором состояния подводных ландшафтов. Для качественной оценки состояния и прогноза ДПК необходимо комплексно рассматривать факторы как биотические – фито- и зообентос, так и абиотические – гранулометрический состав донных отложений, степень консолидации, морфометрические признаки донного рельефа.
Список литературы
1. Блинова Е.И. Морские водоросли-макрофиты и травы Европейской части России. Изд. ВНИРО, 2007. 113 с.
2. Возжинская В.Б. Видовой состав, распределение и фитогеографическая характеристика донной флоры Белого моря // Донная флора и продукция краевых морей СССР. М.: Наука, 1980. С. 29-62.
3. Возжинская В.Б. Донные макрофиты Белого моря. М.: Наука, 1986. 190 с.
4. Игнатов Е.И. Береговые морфосистемы. М.: Маджента, 2004. С. 350.
5. Калугина-Гутник А.А. Фитобентос Черного моря. Киев: Наукова думка, 1975. 250 с.
6. Промысловые водоросли СССР. М.: Пищевая промышленность, 1971. 269 с.
7. Папунов Д.В. Структура и динамика донных природных комплесов береговой зоны Черного и Белого морей: Автореф. дис…канд. геогр. наук. - М., 1991. - 20 с.
8. Папунов Д.В. Сезонная динамика подводных ландшафтов прибрежно-морской зоны Черного и Белого морей // Материалы XV Международной конференции студентов и молодых ученых «Ломоносов-2008». Москва, 2008. С. 92-93.
9. Петров К.М. Комплексное физико-географическое изучение морских мелкводий // Изв. ВГО. 1973. №2. С.108-117.
опубликовано - декабрь 2012 г.
Macrophytobenthos as an indicator of bottom marine seascapes in coastal zone
Dmitry V. Papunov
In this work the main mechanisms of the dynamics of littoral zone underwater landscapes were investigated. It was shown that the population of various types of macrophytobenthos can be used as a main indicator of underwater landscapes state. As one of the examples the test polygons at Black and White sea were proposed. The depth distribution of the season dynamics of the biomass of macrophytes was identified.
Key words: underwater landscapes; indicator of landscape dynamics; phytobenthos of littoral zone of the sea; Black Sea; White Sea; dynamics of underwater landscapes.
Об авторе
Папунов Дмитрий Валерьевич - Papunov Dmitry Valerievich
кандидат географических наук
Федеральное государственное бюджетное учреждение «Государственный океанографический институт имени Н.Н.Зубова» Росгидромета - ФГБУ «ГОИН» (State Oceangraphic Institute)
geopapu@gmail.com
Корреспондентский адрес: Россия, 119034, Москва, Кропоткинский пер., д.6; телефон (499) 246-72-88
ССЫЛКА НА СТАТЬЮ:
Папунов Д.В. Макрофитобентос как индикатор динамики подводных ландшафтов береговой зоны моря // Вопросы современной альгологии. 2012. № 2 (2). URL: http://algology.ru/121
К другим статьям Международной конференции «Экологическая физиология водных фототрофов». I Сабининские чтения
Карта сайта

|
