|
|
|
|
«Экологическая физиология водных фототрофов» I Сабининские чтения 15 ноября - 29 декабря 2012 г. Влияние элементов минерального питания на структуру фитопланктонного сообщества в северо-восточной части Чёрного моря. Экспериментальные исследования
Лифанчук А.В. Anna V. Lifanchuk
Южное отделение Института океанологии им. П.П. Ширшова РАН
УДК 574.55: 574.24
Ключевые слова: фитопланктон; диатомовые водоросли; кокколитофориды; элементы минерального питания; Чёрное море.
Введение Для морских экосистем характерны межгодовые и сезонные изменения. Особенно выраженно они проявляются на уровне фитопланктонного сообщества, где происходит смена лидирующего комплекса. Полевые наблюдения за фитопланктонным сообществом шельфа северо-восточной части Чёрного моря фиксируют эти изменения в системе диатомовые водоросли – кокколитофориды (Паутова и др., 2007; Силкин и др., 2009). Важно понимать закономерности данного процесса для этих двух групп водорослей, так как они играют ключевую роль в регуляции атмосферного СО2 (Volk, Hoffert, 1985; Frankignoulle et al., 1994; Holligan, Robertson, 1996; Smetacek, 1999; Iglesias-Rodríguez et al., 2002). Но объяснить причины межгодовых и сезонных изменений невозможно, используя лишь полевые наблюдения. Дополнительные данные для понимания механизмов регуляции видовой структуры фитопланктонного сообщества дают экспериментальные исследования с природной популяцией фитопланктона. Существенными факторами, определяющими структуру фитопланктона, являются элементы минерального питания. Азот и фосфор являются основными питательными веществами, которые могут ограничить рост фитопланктона в природной среде (Bergstrom et al., 2008; Davey et al., 2008; Persic et al., 2009). Целью данной работы было определить влияние концентраций элементов минерального питания на видовую структуру фитопланктонного сообщества. Для этого были проведены экспериментальные исследования с добавками азота и фосфора в накопительную культуру фитопланктона северо-восточной части Чёрного моря. Материалы и методы С осени 2011 до августа 2012 года проводили эксперименты с накопительной культурой природного фитопланктона. Объектом исследования служила смешанная культура водорослей, полученная на основе природного сообщества фитопланктона. Пробы были собраны с борта научно-исследовательского судна «Ашамба». Отбор проб производили с горизонта 0 м на станции стандартного разреза от Голубой бухты к центру моря. Станция располагалась над глубиной 50 м (район срединного шельфа). Выращивание проводилось в колбах Эрленмейера емкостью 500 мл (объем среды 200 мл) в термолюминостате, где температура среды соответствовала температуре морской воды в месте отбора проб. Идентификацию видов проводили в соответствии с http://www.algaebase.org и http://www.marinespecies.org. Подсчет числа клеток проходил ежедневно на световом микроскопе в счетной камере Ножотта объемом 0,05 мл. Относительная ошибка счета составила 20 %. Биомассу рассчитывали методом «истинного объема» (Киселев, 1969), при этом использовались данные собственных измерений. Опыты ставили с применением метода планирования экспериментов (Максимов, Федоров, 1969), которые позволяют получать уравнения регрессии, отражающие действие выбранных факторов на изучаемый параметр. В экспериментах применяли периодический (накопительный) режим культивирования. Добавку элементов минерального питания (нитратов в форме KNO3 и фосфатов в форме Na2HPO4) производили в начале эксперимента по единой схеме, представленной в таблице №1. Таблица 1. Схема добавления элементов минерального питания в накопительную культуру водорослей
Результаты и обсуждениe Рост численности (биомассы) клеток фитопланктона в накопительной культуре водорослей соответствовал динамике накопления численности (биомассы) в периодической культуре и хорошо аппроксимировался логической кривой (рис. 1-3). Численность (биомассу) клеток водорослей в стационарной фазе роста можно принять в качестве основного параметра для уравнения регрессии (Силкин, Хайлов, 1988).
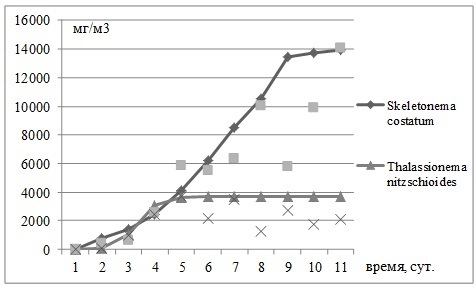
Рис. 1. Рост биомассы Skeletonema costatum и Thalassionema nitzschioides в эксперименте с совместным добавлением нитратов и фосфатов в накопительной культуре водорослей. Опыт проведен 17 июля 2012 г.
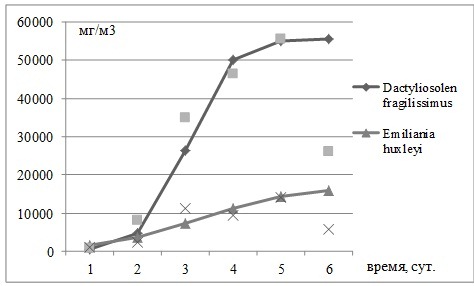
Рис. 2. Рост биомассы Dactyliosolen fragilissimus и Emiliania huxleyi в эксперименте с совместным добавлением нитратов и фосфатов в накопительной культуре водорослей. Опыт проведен 1 июня 2012 г.
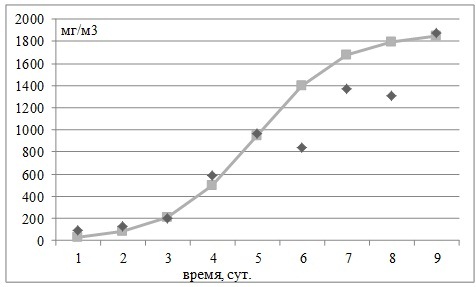
Рис. 3. Рост биомассы Skeletonema costatum в эксперименте с природной популяцией фитопланктона, взятой из поверхностного слоя воды на шельфовой станции в районе Геленджика 1 июня 2012 г. Примечание: в опыте добавлялись фосфор и азот.
При накопительном культивировании природного фитопланктона интенсивное развитие получают виды, которые относятся к доминантам или субдоминантам в природном сообществе. Осень 2011 года характеризуется наличием только диатомового комплекса (табл. 2). Для сентября отмечено положительное влияние одновременной добавки азота и фосфора для таких видов как Leptocylindrus danicus, Chaetoceros curvisrtus, Skeletonema costatum и Dactyliosolen fragilissimus. Для Pseudo-nitzschia pseudodelicatissima добавка азота и фосфора сравнима с ошибкой эксперимента. Это указывает на то, что к этому времени данный вид не получил достаточного развития в природе, и его биомасса в исходной культуре была очень низка. Остальные виды показали интенсивный рост при одновременной добавке элементов минерального питания. В апреле 2012 г. продолжает развиваться Skeletonema costatum и появляется новый вид Thalassionema nitzschioides. Эти виды положительно реагируют на одновременную добавку азота и фосфора. Такие представители диатомового комплекса, как Leptocylindrus danicus, Chaetoceros curvisrtus и Dactyliosolen fragilissimus уходят из фитоцена, их реакция на добавку элементов минерального питания не выходит за пределы доверительного интервала значимости. Диатомея Pseudo-nitzschia pseudodelicatissima получает преимущественное развитие при добавке азота. Всё это указывает на то, что в период с сентября по октябрь снижаются концентрации азота и фосфора в среде. Виды Leptocylindrus danicus, Chaetoceros curvisrtus и Dactyliosolen fragilissimus приурочены к высоким концентрациям элементов минерального питания, а Pseudo-nitzschia pseudodelicatissima, Skeletonema costatum и Thalassionema nitzschioides могут развиваться при относительно низких концентрациях азота и фосфора.
Таблица 2. Уравнения регрессии для биомассы (Wst) доминирующих видов микроводорослей в стационарной фазе роста накопительной фитопланктона, взятой из поверхностного слоя воды на шельфовой станции в районе г. Геленджика осенью 2011 г.
Весной 2012 года доминирующими видами в экспериментах были Pseudo-nitzschia pseudodelicatissima и Skeletonema costatum (табл. 3). В марте диатомовые водоросли не зависели от добавок азота и фосфора в накопительную культуру. За рост биомассы диатомей был ответственен другой фактор, предположительно кремний. Но уже в апреле ситуация изменилась – Pseudo-nitzschia pseudodelicatissima стала лимитирована по азоту и фосфору. Добавки этих элементов минерального питания не влияли на рост биомассы Skeletonema costatum. Вероятно, что в апреле месяце с усилившимся речным стоком происходит интенсивное поступление кремния в прибрежную часть Чёрного моря, поэтому данный элемент перестает быть лимитирующим.
Таблица 3. Уравнения регрессии для биомассы (Wst) доминирующих видов микроводорослей в стационарной фазе роста накопительной фитопланктона, взятой из поверхностного слоя воды на шельфовой станции в районе г. Геленджика весной 2012 г.
В летний период 2012 года кокколитофориды доминировали в фитопланктонном сообществе. Численность Emiliania huxleyi в июне 2012 г. достигала 9 х 106 кл/л. Доля диатомей в структуре фитопланктонного сообщества в это время была невелика, Диатомеи были представлены Chaetoceros curvisrtus, Skeletonema costatum, Thalassionema nitzschioides и Dactyliosolen fragilissimus. Таким образом, Emiliania huxleyi представляла собой монокультуру с редкими вкраплениями диатомей в сообществе фитопланктона, и была лимитирована по азоту и фосфору (табл. 4). В июле происходит смена лидирующего комплекса в структуре фитопланктонного сообщества. В системе диатомовые водоросли – кокколитофориды лидируют диатомеи, за рост биомассы (численности) которых ответственны оба элемента минерального питания. Величины коэффициентов уравнения регрессии, описывающих уровень накопленной биомассы, указывают на то, что в середине лета наиболее значимым фактором для Emiliania huxleyi. являлась концентрация фосфатов в накопительной культуре. Считалось, что кокколитофориды способны активно развиваться при низких концентрациях фосфора и, следовательно, при высоких соотношениях азота и фосфора (Egge, Aksness, 1992; Egge, Hiemdall, 1994). Но как оказалось, низкое соотношение азота и фосфора в среде не всегда указывает на азот как на фактор, лимитирующий развитие кокколитофорид. Установлено, что основным фактором, определяющим уровень количественного развития кокколитофориды Emiliania huxleyi на шельфе северо-восточной части Черного моря, является концентрация фосфора (Силкин и др., 2009). Таким образом, эксперименты с накопительной культурой водорослей, проведенные летом 2012 г., подтверждают данную гипотезу.
Таблица 4. Уравнения регрессии для биомассы (Wst) доминирующих видов микроводорослей в стационарной фазе роста накопительной фитопланктона, взятой из поверхностного слоя воды на шельфовой станции в районе г. Геленджика летом 2012 г.
Таким образом, в марте–апреле 2012 г. во время усиленной стратификации водной толщи Чёрного моря образовалась зона пониженной концентрации азота. Вследствие этого не происходило развитие диатомовых водорослей ранней весной. В июле 2012 г. усилился обмен минеральными веществами в период более интенсивного вертикального перемешивания, что привело к доминированию диатомей в структуре фитопланктонного сообщества. Выводы Осенью 2011 г. и весной 2012 г. доминировали диатомовые водоросли. В осенний период снизилась концентрация элементов минерального питания в среде, что привело к доминированию видов, хорошо развивающихся при низких концентрациях азота и фосфора. В марте диатомеи не зависели от концентраций азота и фосфора в накопительной культуре, предположительно, они были лимитированы по кремнию. В апреле происходило поступление этого элемента с речными стоками, и развитие диатомовых водорослей стало зависеть от совместной добавки азота и фосфора. Летом 2012 г. доминантами являлись кокколитофориды. Повышение концентрации фосфора в морской воде при относительно низких концентрациях азота ответственно за увеличение численности кокколитофорид и преобладание этих водорослей в структуре сообщества. Совместное повышение концентраций азота и фосфора приводило к доминированию диатомовых водорослей и повышению уровня трофности прибрежных вод. Результаты данных исследований убедительно показывают, что изменяя концентрации азота и фосфора, можно существенно разделить условия доминирования в паре диатомовые-кокколитофориды.
Список литературы 1. Киселев И.А. Планктон морей и континентальных водоемов // Вводные и общие вопросы планктонологии. Л.: Наука, 1969. Т.1. 657 с. 2. Максимов В.Н., Федоров В.Д.. Применение методов математического планирования эксперимента при отыскании оптимальных условий культивирования организмов. М.: Изд-во МГУ, 1969. 128 с. 3. Паутова Л.А., Микаэлян А.С., Силкин В.А. Структура планктонных фитоценов шельфовых вод северо-восточной части Черного моря в период массового развития Emiliania huxleyi в 2002-2005 гг. // Океанология. 2007. Т. 47. №3. С. 408-417. 4. Силкин В.А., Паутова Л.А., Микаэлян А.С. Рост кокколитофориды Emiliania huxleyi (Lohmann) Hay et Mohler в северо-восточной части Черного моря, лимитированный фосфором // Альгология. 2009. V. 19. №2. С. 135-143. 5. Силкин В.А., Хайлов К.М. Биоэкологические механизмы в аквакультуре. Л.: Наука, 1988. 230 с. 6. Bergstrom, A. K., Jonsson, A., Jansson, M. Phytoplankton responses to nitrogen and phosphorus enrichment in unproductive swedish lakes along a gradient of atmospheric nitrogen deposition // Aquatic Biology. 2008. 4 (1). P. 55-64. 7. Davey, M., Tarran, G. A., Mills, M. M., Ridame, C., Geider, R. J., LaRoche, J. Nutrient limitation of picophytoplankton photosynthesis and growth in the tropical north atlantic // Limnology and Oceanography. 2008. 53 (5). P. 1722-1733. 8. Egge J.K., Aksness D.L. Silicate as regulating nutrient in phytoplankton competition // Mar. Ecol. Prog. Ser. 1992. 83. P. 281-289. 9. Egge J.K., Hiemdall B.R. Blooms of phytoplankton including Emiliania huxleyi (Haptophyta). Effects of nutrient supply in different N:P ratios // Sarsia. 1994. 79. P. 333-348. 10. Frankignoulle M, Canon C, Gattuso J-P. Marine calcification as a source of carbon dioxide: Positive feedback of increasing atmospheric CO2 // Limnol Oceanog. 1994. 39. P. 458–462. 11. Holligan PM, Robertson JE. Significance of ocean carbonate budgets for the global carbon cycle // Glob Change Biol. 1996. 2. P. 85–95. 12. Iglesias-Rodríguez MD, et al. Representing key phytoplankton functional groups in ocean carbon cycle models: Coccolithophorids // Glob Biogeochem Cycles. 2002. 16. P. 1100. DOI: 10.1029/2001GB001454. 13. Persic, V., Horvatic, J., Has-Schon, E., Bogut. Changes in n and p limitation induced by water level fluctuations in nature park kopacki rit (croatia): nutrient enrichment bioassay // Aquatic Ecology. 2009. 43 (1). P. 27-36. 14. Smetacek V. Diatoms and the ocean carbon cycle // Protist. 1999. 150. P. 25–32. 15. Volk T, Hoffert M.I. The Carbon Cycle and Atmospheric CO2 / Natural Variations Archean to Present. Eds Sundquist ET, Broecker WS. Washington, DC: American Geophysical Union. 1985. P. 99–110. опубликовано - декабрь 2012 г.
The influence of mineral elements on the structure of the phytoplankton community in the north-east Black sea. Experimental studies Anna V. Lifanchuk The influence of the concentration of mineral elements on the species composition of the phytoplankton community in the system of diatom-coccolithophorids in 2011-2012 was revealed. Increasing the concentration of phosphorus in seawater at relatively low concentrations nitrogen is responsible for the increase in the number of coccolithophorids and the predominance of these algae in the community. The joint increasing concentrations of nitrogen and phosphorus led to the dominance of diatoms. Key words: phytoplankton; diatom; coccolithophorids; mineral elements; Black sea.
Об авторе Лифанчук Анна Викторовна - Lifanchuk Anna Viktorovna младший научный сотрудник lifanchuk.anna@mail.ru Корреспондентский адрес: Россия, 353470, Краснодарский край, г. Геленджик, ул. Просторная 1-г. Телефон/факс 8-861-41-280-89.
ССЫЛКА НА СТАТЬЮ: Лифанчук А.В. Влияние элементов минерального питания на структуру фитопланктонного сообщества в северо-восточной части Чёрного моря. Экспериментальные исследования // Вопросы современной альгологии. 2012. № 2 (2). URL: http://algology.ru/120

|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||