|
|
|
Влияние океанических концентраций серы на рост диатомей в северо-восточной части Черного моря
Лифанчук А.В., Федоров А.В. Anna V. Lifanchuk, Aleksey V. Fedorov
Южное отделение Института океанологии им. П.П. Ширшова РАН (г. Геленджик)
УДК 581.1:574.522
В работе проверяется следующая гипотеза: повышение концентрации серы до океанического уровня способствовало таксономическому разнообразию фитопланктона. Поскольку черноморский фитопланктон адаптирован к концентрациям серы, равным половине океанической, повышение концентрации этого элемента до океанического уровня не должно приводить к снижению продукционных свойств доминирующих видов. В экспериментальных условиях исследовалось влияние повышений концентраций азота, фосфора и серы на концентрацию биомассы диатомей Leptocylindrus danicus и Chaetoceros sp. в стационарной фазе накопительной культуры. Показано, что повышение концентрации серы приводит к существенному снижению продукционных свойств диатомей. Следовательно, на уровне центрических диатомей предложенная гипотеза не может быть принята. Ключевые слова: фитопланктон; диатомеи; азот; фосфор; сера; Черное море; океан.
Введение Основными доминантами фитопланктона современного океана являются диатомовые, динофлагелляты и кокколитофориды (Falkowski et al., 2004). Считается, что диатомеи распространились в океане около 250 миллионов лет назад, история кокколитофорид начинается на 30 млн. лет позднее, а динофлагелляты стали доминировать в океане примерно 200 млн лет назад (Katz et al., 2004). Кроме того, в океане динофлагелляты демонстрируют более высокое видовое разнообразие по сравнению с пресными водами. Существует точка зрения, что доминированию этих трех таксономических групп способствовало увеличение концентрации сульфатов в океане (Ratti et al., 2011). В Черном море природная популяция фитопланктона адаптирована к солености, равной половине океанической. Также, концентрация серы составляет половину океанической. По своему таксономическому составу фитопланктон является типично морским (Pautova et al., 2011). Следовательно, согласно этой гипотезе, следует ожидать, что повышение концентрации серы должно повышать продукционные свойства существующих доминантов или, по крайней мере, не будет иметь значительных последствий для продуктивности фитопланктона Черного моря. Основной целью настоящего исследования была проверка справедливости следующей гипотезы: повышение концентрации серы до океанического уровня не приводит к снижению продукционных характеристик представителей черноморского фитопланктона. Для этого в экспериментальных условиях исследовалось влияние повышенной концентрации серы на рост диатомей природной популяции черноморского фитопланктона.
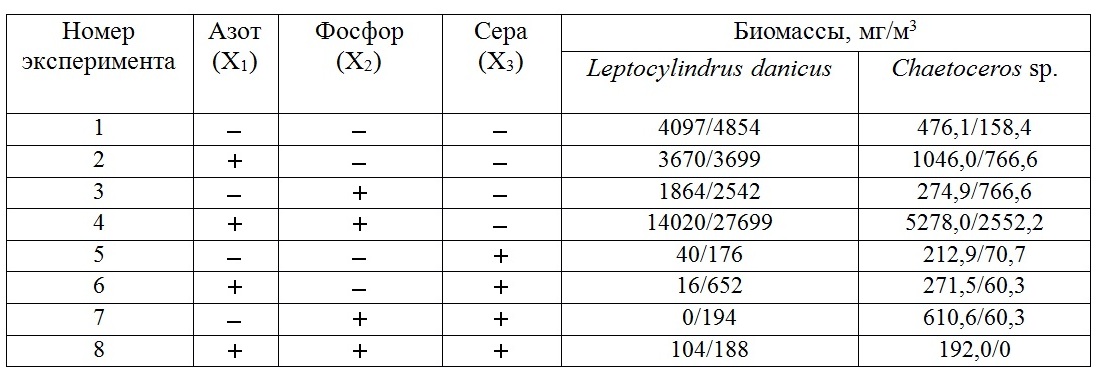
Методы Экспериментальные исследования. В июне 2015 г. в береговой лаборатории ЮО ИО РАН (г. Геленджик) проводились исследования по влиянию добавок нитратов, фосфатов и сульфатов на структуру фитопланктонного сообщества. Вода для экспериментов была взята с поверхности (0–0,5 м) на шельфовой станции стандартного разреза (глубина 50 м). Пробы воды отбирали в пластиковые емкости объемом 5 л с одновременной фильтрацией через два слоя мельничного газа №36 для удаления зоопланктона. Для определения структуры сообщества природного фитопланктона в начале эксперимента 2 л нефильтрованной воды сконцентрировали при помощи камеры обратной фильтрации. Эксперименты проводили в 0,5-литровых колбах Эрленмейера, объем культуральной среды составлял 200 мл. Конечная концентрация элементов минерального питания в колбе после добавления составляла 12–14 мкМ, 1 мкМ и 14 мМ для нитратов, фосфатов и серы соответственно. Для эксперимента использовалась схема полного факторного эксперимента 32 (таблица 1) (Максимов, 1980). В Таблице 1 знак «-» указывает, что добавление элементов минерального питания не производили, а «+» означает, что добавление было выполнено. Для вычисления влияния элементов минерального питания на увеличение биомассы или численности в стационарной фазе роста, знак «-» или «+» должен быть заменен на X1 (добавка азота), X2 (добавка фосфора) и X3 (добавка серы) в уравнение регрессии: Y = bo + b1X1 + b2X2 + b3X3 + b12X1X2 + b13X1X3 + b23X2X3 + b123X1X2X3, где коэффициент bo равен среднему арифметическому результатов эксперимента (биомасс) во всех вариантах; b1X1 – результаты эксперимента в вариантах с добавкой азота; b2X2 – результаты эксперимента в вариантах с добавкой фосфора; b3X3 – результаты эксперимента в вариантах с добавкой серы; b12X1X2 – результаты эксперимента в вариантах с совместной добавкой азота и фосфора; b13X1X3 – результаты эксперимента в вариантах с совместной добавкой азота и серы; b23X2X3 – результаты эксперимента в вариантах с совместной добавкой фосфора и серы; b123X1X2X3 – результаты эксперимента в вариантах с совместной добавкой азота, фосфора и серы. В Таблице 1 приведены значения биомасс для диатомей Leptocylindrus danicus и Chaetoceros sp. в двух повторностях эксперимента с добавками элементов минерального питания в накопительную культуру природного фитопланктона.
Таблица 1. Схема эксперимента для изучения влияния нитратов, фосфатов и добавок серы на максимальную биомассу фитопланктона
В эксперименте использовали аппарат для культивирования, где температура среды соответствовала температуре морской воды в месте отбора проб. Интенсивность падающего света поддерживалась на уровне 58–61 мкмоль/м2 ФАР; фотопериодический цикл составлял 16 ч света : 8 ч темноты. Во всех экспериментах применялся периодический режим культивирования. Подсчет количества клеток водорослей проводили ежедневно, используя камеру Ножжота 0,05 мл сразу же после отбора проб культуры. Для расчета биомассы использовался метод, основанный на геометрической форме клеток (Киселев, 1969; Hillebrand et al., 1999). Биомасса была выражена в единицах сырого веса (мг/м3).
Результаты экспериментальных исследований Влияние азота, фосфора и серы на рост диатомеи Leptocylindrus danicus Уравнение регрессии, описывающее влияние повышения концентрации азота, фосфора и серы на рост мелкоклеточной диатомеи, имеет вид: Wst = 3996 + 2273X1 + 1844X2 – 3823X3+ 2416X1X2 + 2206X1X3 – 1894,5X2X3 – 2459X1X2X3 (95% доверительный интервал – 2844). В отсутствии добавки серы (варианты 1–4, табл. 1) влияние повышенных концентраций азота и фосфора описывается следующим регрессионным уравнением: Wst = 7815,5 + 4479X1 + 3738X2 + 4874,5X1X2 (95% доверительный интервал – 5413). Из уравнения следует, что только одновременная добавка азота и фосфора стимулирует рост диатомеи. Добавление одного только элемента не приводит к усилению роста диатомеи. Добавление серы (варианты 5–8, табл. 1) приводит к резкому падению продуктивности диатомеи: Wst = 173 + 68X1 – 51X2 – 43X1X2 (95% доверительный интервал – 273). В этом случае влияние азота и фосфора существенно ниже ошибки эксперимента. Совместное влияние повышенных концентраций азота и серы существенно зависит от концентрации фосфора. Если фосфор находится на верхнем уровне, то добавление серы существенно не влияет в случае, если выращивание происходит на природной воде без добавления азота: Wst = 5839 + 4689X1 – 5717X3 – 4664X1X3 (95% доверительный интервал – 5405). Добавление азота приводит к тому, что повышение концентрации серы до уровня океанической приводит к существенному падению продукционных свойств данной диатомеи. При культивировании на природной воде без добавления фосфора продуктивность в среднем падает более чем в два раза, а реакция водоросли на повышение концентрации серы зависит от уровня концентрации азота, но в любом случае эффект повышения концентрации серы значим: Wst = 2152 – 142X1 – 1928X3 – 253X1X3 (95% доверительный интервал – 398). Рассмотрение взаимодействия пары фосфор и сера показывает, что отрицательный эффект зависит от уровня концентрации азота в среде. Если азот находится на верхнем уровне, то уравнение имеет вид: Wst = 6269 + 4259X2 – 6029X3 – 4353X2X3 (95% доверительный интервал – 5404). В этом случае, если фосфор не добавляется, существенного влияния повышения концентрации серы не наблюдается. Если фосфор находится на верхнем уровне, то повышение концентрации серы до океанического уровня негативно влияет на скорость роста диатомеи. В отсутствии добавки азота продуктивность значительно падает, а реакция водоросли на изменение концентрации серы зависит от концентрации фосфора в среде: Wst = 1722 – 572X2 – 1617X3 + 564X2X3 (95% доверительный интервал – 415). При культивировании на природной воде без добавления фосфора эффект повышения концентрации серы очень значительный. При добавлении фосфора отрицательный эффект повышения концентрации серы снижается. Влияние азота, фосфора и серы на рост диатомеи Chaetoceros sp. Уравнение регрессии, описывающее влияние повышения концентрации азота, фосфора и серы на рост диатомеи Chaetoceros sp., имеет вид: Wst = 800 + 942X1 + 834X2 – 777X3+ 635X1X2 – 1050,5X1X3 – 772X2X3 – 767X1X2X3 (95% доверительный интервал – 599,5). В вариантах без добавления серы (варианты 1–4, табл. 1) только совместное добавление азота и фосфора повышает биомассу этой диатомеи, отдельное повышение концентрации одного из элементов не приводит к существенному росту биомассы: Wst = 1415+ 996X1 + 803X2 + 701X1X2 (95% доверительный интервал – 1114). Добавление серы снижает рост биомассы более чем в 7 раз, при этом влияние повышения концентраций азота и фосфора сравнимо с ошибкой эксперимента: Wst = 185 – 54X1 + 31X2 – 66X1X2 (95% доверительный интервал – 253). Влияние концентрации азота и серы различно и зависит от концентрации фосфора. При повышенной концентрации фосфора (варианты 3, 4, 7, 8, табл. 1) максимальный отрицательный эффект повышения концентрации серы достигается при добавлении азота, при выращивании же в воде без добавления азота этот эффект близок к ошибке эксперимента: Wst = 1217 + 789X1 – 1001X3 - 908X1X3 (95% доверительный интервал – 1125). В вариантах без добавления фосфора (варианты 1, 2, 5, 6, табл. 1) биомасса резко снижается и отрицательный эффект повышения концентрация серы существенно не зависит от уровня концентрации азота: Wst = 383 + 153X1 – 229X3 – 141X1X3 (95% доверительный интервал – 196,5). Влияние пары фосфор и сера существенно зависит от концентрации азота в среде. При повышенной концентрации азота (варианты 2, 4, 6, 8, табл. 1) отрицательный эффект добавления серы максимален при нахождении фосфора на верхнем уровне и минимален, когда фосфор не добавляется: Wst = 1271 + 735X2 – 1140X3 - 770X2X3 (95% доверительный интервал – 1095). Культивирование Chaetoceros sp. на природной воде без добавления азота (варианты 1, 3, 5, 7, табл. 1) снижает биомассу практически в 4 раза, эффект добавки как фосфора, так и серы не влияет на уровень биомассы: Wst = 329 + 99X2 – 90X3 – 2,5X2X3 (95% доверительный интервал – 324).
Обсуждение В монокультурах водорослей было показано, что отношение азота и фосфора для каждого вида является характерным параметром (Rhee, 1978; Rhee, Gotham, 1981; Силкин, Хайлов, 1988). Оптимальное соотношение азота и фосфора в биомассе изменяется в широких пределах (Klausmeier et al, 2004). Было показано, что для диатомовых водорослей оптимальное соотношение азота и фосфора близко или выше соотношения Редфилда (Silkin et al., 2014). Для культур одноклеточных водорослей также имеет значение соотношение исходных концентраций этих элементов в окружающей среде. Это соотношение может быть либо не оптимальным, либо оптимальным для интенсивного роста. Оптимальное соотношение приблизительно равно отношению минимальных квот (Силкин, Хайлов, 1988). Очевидно, что влиять на рост водорослей, помимо азота и фосфора, может и такое эссенциальное вещество как сера, которая является структурным элементом аминокислот и, следовательно, связана с азотным обменом. Учитывая, что при делении клетки происходит синтез ДНК и РНК, для которых необходим фосфор, этот фактор также важен для адаптации клеток к повышенным концентрациям серы. В наших исследованиях концентрация кремния была высокой (> 4 µM в большинстве случаев) и не могла ограничить рост диатомовых водорослей. Наблюдаемые концентрации кремния были намного выше, чем константа полунасыщения для поглощения кремния (Krause et al., 2012). В наших экспериментах в отсутствии добавки серы повышение концентрации только одного элемента питания (азота или фосфора) не приводило к существенному росту Leptocylindrus danicus и Chaetoceros sp. Но оба вида демонстрируют интенсивный рост при одновременной добавке азота и фосфора. Учитывая, что отношение азота к фосфору в добавках было равно 14, т.е. близко к соотношению Редфилда, можно ожидать, что такое соотношение будет оптимальным для этих водорослей. Повышение концентрации серы до океанического уровня приводит к существенному ингибированию роста обоих видов диатомей. В этом случае добавление азота и фосфора не приводит к заметному росту диатомей. Данные виды распространены в Северной Атлантике (Barton et al., 2013) и, следовательно, хорошо адаптированы к океаническим концентрациям серы. В северо-восточной части Черного моря они также входят в число доминантов и демонстрируют интенсивное развитие (Pautova et al., 2007; Silkin et al., 2013), что говорит об их хорошей адаптации к существующей концентрации серы, равной половине океанической. Отрицательная реакция черноморских видов диатомей на повышение концентрации серы указывает на то, что эти виды в процессе длительной эволюции в условиях пониженной концентрации серы утратили способность быстро адаптироваться к ее повышенным концентрациям. Таким образом, повышение концентрации серы до океанического уровня приводит к ингибированию роста центрических диатомей черноморского фитопланктона, адаптированного к концентрациям серы, равным половине океанической. Следовательно, повышение концентрации серы не могло стимулировать таксономическое разнообразие диатомового комплекса. Работа выполнена в рамках госзадания № 0149-2014-0056.
Список литературы 1. Киселев И.А. Планктон морей и континентальных водоемов. Т.I. Вводные и общие вопросы планктологии. – Л.: Наука Ленингр. отд., 1969. – 658 с. 2. Максимов В.Н. Многофакторный эксперимент в биологии. – М.: Издательство Московского университета, 1980. – С. 164–190. 3. Силкин В.А., Хайлов К.М. Биоэкологические механизмы управления в аквакультуре. – Ленинград: Наука, 1988 – 230 с. 4. Barton A.D., Finkel Z.V., Ben B., Ward A., David A.C., Johns D.G., Michael J. On the roles of cell size and trophic strategy in North Atlantic diatom and dinoflagellate communities // Limnology and Oceanography. 2013. 58(1). P. 254–266. 5. Falkowski P.G., Katz M.E., Knoll A.H., Quigg A., Raven J.A., Schofield O., Taylor F.J.R. The evolution of modern eukaryotic phytoplankton // Science. 2004. 305. P. 354–360. 6. Hillebrand H, Durselen C, Kirschtel D, Pollingher U., Zohary T. Biovolume calculation for pelagic and benthic microalgae // J Phycol. 1999. 35. P. 403–424. 7. Katz M.E., Finkel Z.V., Grzebyk D., Knoll A.H., Falkowski P.G. Evolutionary trajectories and biogeochemical impacts of marine eukaryotic phytoplankton // Annual Review of Ecology Evolution and Systematics. 2004. 35. P. 523–556. 8. Klausmeier C.A., Litchman E., Daufresne T., Levin S.A. Optimal nitrogen-to-phosphorus stoichiometry of phytoplankton // Nature. 2004. 429. P. 171–174. 9. Krause J.W., Brzezinski M.A., Villareal T.A., Wilson C. Increased kinetic efficiency for silicic acid uptake as a driver of summer diatom blooms in the North Pacific subtropical gyre // Limnology and Oceanography. 2012. 57. P. 1084–1098. 10. Pautova L.A., Mikaelyan A.S., Silkin V.A. The structure of plankton community in shelf waters of north-eastern part of the Black Sea in the period of mass bloom of Emiliania huxleyi in 2002–2005 // Oceanology. 2007. 47. P. 408–417. 11. Ratti S., Knoll A.H., Giordano M. Did sulfate availability facilitate the evolutionary expansion of chlorophyll a plus c phytoplankton in the oceans? // Geobiology. 2011. 9. P. 301–312. 12. Rhee G.-Y. Effects of N:P atomic ratios and nitrate limitation on algal growth, cell composition and nitrate uptake // Limnology and Oceanography. 1978. 23. P. 10–25. 13. Rhee G.-Y., Gotham I.J. The effect of environmental factors on phytoplankton growth: light and the interactions of light with nitrate limitation // Limnology and Oceanography. 1981. 26. P. 649–659. 14. Silkin V.A., Pautova L.A., Lifanchuk A.V. Physiological Regulatory Mechanisms of the Marine Phytoplankton Community Structure // Russian Journal of Plant Physiology. 2013. V.60, №4. P. 541–548. Статья поступила в редакцию 10.11.2017
Influence of the oceanic concentration of the sulphur on diatoms growth in the NE Black Sea Anna V. Lifanchuk, Aleksey V. Fedorov Southern Branch of the P.P. Shirshov Institute of Oceanology of RAS (Gelendzhik, Russia) The following hypothesis is tested in the paper: increasing the sulfur concentration to the oceanic level has contributed to the taxonomic diversity of phytoplankton. Since the Black Sea phytoplankton is adapted to sulfur concentrations equal to half the oceanic level, increasing the concentration of this element to the oceanic level should not lead to a decrease in the production properties of the dominant species. In the experimental conditions, the effect of increasing concentrations of nitrogen, phosphorus and sulfur on the biomass concentration of diatom Leptocylindrus danicus and Chaetoceros sp. in the stationary phase of a bath culture. It is shown that an increase in the sulfur concentration leads to a significant decrease in the production properties of diatoms. Consequently, at the level of centric diatoms, the proposed hypothesis c’ not be accepted. Key words: phytoplankton; diatoms; nitrogen; phosphorus; sulfur; Black Sea; ocean.
Об авторах Лифанчук Анна Викторовна − Lifanchuk Anna Viktorovna младший научный сотрудник, Южное отделение ФГБУН «Институт океанологии им.П.П.Ширшова РАН», Геленджик, Россия (Southern branch of the P.P. Shirshov Institute of Oceanology RAS, Gelendzhik, Russia)
Фёдоров Алексей Владимирович − Fedorov Aleksey Vladimirovich инженер-исследователь, Южное отделение ФГБУН «Институт океанологии им.П.П.Ширшова РАН», Геленджик, Россия (Southern branch of the P.P. Shirshov Institute of Oceanology RAS, Gelendzhik, Russia)
Корреспондентский адрес: Россия, 353470, Краснодарский край, г. Геленджик, ул. Просторная 1-г. Телефон/факс 8-861-41-280-89.
ССЫЛКА НА СТАТЬЮ: Лифанчук А.В., Федоров А.В. Влияние океанических концентраций серы на рост диатомей в северо-восточной части Черного моря // Вопросы современной альгологии. 2017. № 2 (14). URL: http://algology.ru/1179 Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно.
При перепечатке ссылка на сайт обязательна
На ГЛАВНУЮ
|
|||
|
| ||