|
|
|
|
«Экологическая физиология водных фототрофов» I Сабининские чтения 15 ноября - 29 декабря 2012 г. К вопросу о влиянии структуры биотопа на динамику зарослей черноморской грациллярии
Переладов М.В. Mikhail V. Pereladov
Всероссийский научно-исследовательский институт рыбного хозяйства и океанографии
УДК 582.273
На основании водолазных наблюдений с 1981 по 2008 гг. рассматривается влияние штормового переноса рыхлых грунтов (песка и гальки) на структуру прибрежных биотопов и, в частности, на динамику зарослей черноморской грациллярии. Ключевые слова: Черное море; грациллярия; биотоп; динамика.
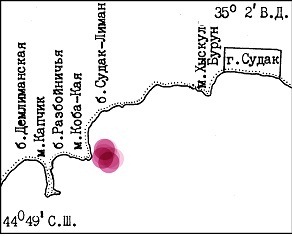
Введение Красные водоросли рода Gracillaria – широко распространенная группа видов, обитающая во многих морях Мирового Океана. Интерес к этим водорослям определяется высоким содержанием в них ценного полисахарида – агара, широко используемого в химической, пищевой и микробиологической промышленности. Во многих странах (преимущественно Юго-Восточной Азии) грациллярию успешно культивируют в условиях марикультуры, что позволяет получать из этих водорослей высококачественную товарную продукцию, пользующуюся постоянным спросом на мировом рынке (Вилкова, 2008; Вилкова и др., 2008). В России промышленного культивирования грациллярии в настоящий момент нет. По своим экологическим предпочтениям водоросли рода Gracillaria можно разделить на две группы – прикрепленные и неприкрепленные. Систематическое положение видов, входящих в эти группы постоянно пересматривается различными исследователями, поэтому в данной работе будет использоваться термин «форма». Род Gracillaria в Черном море представлен двумя формами – прикрепленная, приуроченная к ракушечникам прибрежных акваторий открытого моря (G. verrucosa), и неприкрепленная, приуроченная к зарослям морских трав в лиманах и заливах (G. dura), (Калугина, Миронова, 1985; Миронова, 2010). Несмотря на обилие работ, посвященных грациллярии (Миронова, 2002; 2005; Блинова 2007), экологические предпочтения этого вида изучены слабо. В литературе крайне мало данных, описывающих особенности взаимодействия грациллярии с другими компонентами биоценоза, роли абиотических условий в развитии и многолетней динамике ее зарослей. Особенно это касается прикрепленной формы грациллярии, образующей нестабильные во времени и пространстве поселения в прибрежной зоне моря. Исходя из этих предпосылок, задачи настоящей работы заключались в поиске и описании состояния естественных зарослей грациллярии на акватории Черного моря и в выяснении причин, определяющих многолетнюю динамику ее зарослей. Материал и методики Материалом исследования послужили данные, собранные в ходе водолазных гидробиологических съемок, проводившихся в 1981-1994 гг. и 2005-2008 гг. на акватории Черного моря. Весь материал собран непосредственно автором с использованием легководолазного снаряжения в диапазоне глубин от уреза воды до 40 м по стандартным гидробиологическим методикам (Блинова и др., 2005). В ходе водолазных съемок 1981-1994 гг. планомерно обследовались два участка прибрежной акватории: Судакский залив в Крыму и акватория, прилегающая к мысу Большой Утриш на Кавказе. На этих участках проводились регулярные многолетние наблюдения за динамикой бентосных сообществ и, в частности, за состоянием зарослей грациллярии. В 2005-2008 гг. было продолжено изучение зарослей грациллярии, на тех же участках акватории Черного моря. В ходе каждой водолазной съемки на каждом участке выполнялось несколько гидробиологических разрезов от уреза воды до глубины 15-20 м. На каждом разрезе фиксировалось наличие известных по литературным и собственным данным биотопов, пригодных доля обитания различных форм грациллярии. При нахождении зарослей грациллярии, в соответствующем биотопе закладывалось несколько учетных площадок площадью 1 м2, на которых производился тотальный сбор всех талломов. При малой плотности зарослей сбор талломов грациллярии производится на учетной полосе шириной 1 м и протяженностью не менее 10 м. Учетные полосы закладывались вдоль изобат. Камеральная обработка проб включала в себя фотографирование отдельных талломов грациллярии и измерение их сырого веса. Результаты В период с 1981 по 1994 гг. изучение собственно грациллярии не входило в спектр работ Крымской экспедиции ВНИРО (КРЭКС), поэтому все данные о распределении и многолетней динамике ее зарослей были собраны попутно, в ходе бентосных съемок, направленных на изучение воздействия промышленного культивирования моллюсков на экосистему прибрежной зоны Черного моря. На обоих полигонах заросли прикрепленной формы грациллярии были найдены в типичных для этой формы биотопах – на галечниках, расположенных на глубине 10-15 м. На акватории Судакского залива галечник с грациллярией был обнаружен к западу от мыса Коба-Кая, замыкающего с юга бухту Судак-Лиман (Рис. 1).
Рис. 1. Среднемноголетнее расположение зарослей грациллярии (форма Gracillaria verrucosa) на акватории Судакского залива Черного моря.
Подводный береговой склон на этом участке акватории бухты в диапазоне глубин от уреза воды до 6-10 м представляет собой осыпь крупных глыб и отдельных участков монолитной скалы со средним наклоном 30-45о. Вдоль подножия глыб и скал простирается пояс отдельных глыб, валунов разной размерности и мидийного ракушечника шириной 10-15 м, на 10-12 м переходящий в галечник с отдельными валунами. Ширина галечного пояса не превышает 100 м и на глубине 16-18 м он замещается песком. Вдоль берега галечник простирается на 150-200 м, ограниченный с юга полем песка, а с севера – зарослями морской травы зостеры, произрастающей на смешанном песчано-галечном грунте. Заросли грациллярии в этом районе были приурочены к центральной части описанного галечника и занимали в разные годы от 1000 до 5000 м2. Гранулометрический состав грунта биотопа, в котором были обнаружены талломы прикрепленной грациллярии, представлял собой смесь собственно гальки (50%), крупнозернистого песка (20%) и иловых фракций (20%). В незначительных количествах в грунте встречались мелкие валуны и ракушечник (10%). В зообентосе биоценоза галечников доминирующими видами были двустворчатый моллюск Modiola adriatica и седентарные полихеты рода Terebellida, второстепенными видами – двустворчатый моллюск Mytilaster linneatus и ряд мелких гастропод. Особи модиолы были отмечены непосредственно среди гальки, полупогруженные в грунт. Плотность модиол достигала 150-180 экз./м2 при биомассе порядка 200 г/м2. Особи митиллястера и теребеллид встречались на валунах, лежащих на поверхности грунта, и плотность их поселений существенно варьировала в зависимости от наличия крупнообломочного материала. В фитоценозе галечников по биомассе доминировали кодиум (Codium vermilara) и филлофора (Phyllophora nervosa), талломы которых располагались на крупнообломочном материале и Nereia filiformis, приуроченная к гальке и ракушечнику на поверхности грунта. Талломы грациллярии были приурочены к мелкой гальке, погруженной в рыхлый грунт на 1-3 см.
Фото 1-3. Талломы грациллярии (Gracillaria verrucosa), собранные в биотопе галечников. Судакский залив Черного моря, глубина 14 м, июль 2008 года.
Плотность талломов грациллярии в биотопе галечников в районе поселка Новый Свет за весь период исследования с 1981 по 2008 гг. варьировала от нуля до 40-50 экз./м2. В этих же районах неоднократно были отмечены талломы грациллярии на раковинах брюхоногого моллюска рапаны (Rapana venosa), причем как на неподвижных, зарывшихся в грунт моллюсках, встреченных среди галечников, так и на подвижных моллюсках, отмеченных на песчаном грунте в ближайших окрестностях от галечников.
Фото 4-5. Талломы грациллярии (Gracillaria verrucosa), на раковинах Rapana venosa . Судакский залив Черного моря, глубина 10-18 м, июль 2008 года.
В районе мыса Большой Утриш грациллярия была найдена на галечнике, расположенном на северо-западном склоне острова (Рис. 2).
Рис. 2. Карта-схема расположения зарослей грациллярии в районе мыса Большой Утриш, Черное море
Подводный береговой склон в районе мыса Большой Утриш в диапазоне глубин от уреза воды до 3-4 м представляет собой слабо наклоненное продолжение галечно-валунного пляжа шириной 30-50 м. В этом диапазоне глубин подводный склон занимают заросли макрофитов с доминированием цистозиры (Cysroseira crinita и C. barbata). На глубине 4-6 м, склон резко обрывается глыбово-валунной осыпью до глубины 10-12 м и переходит в плоское галечное плато шириной 10-20 м. Глубже начинается глыбовая осыпь, доходящая до глубины 20-30 м и переходящая затем в заиленный песок. На обеих осыпях в составе макрофитов доминирует филлофора и кодиум. Протяженность среднего галечного плато вдоль берега составляет порядка 100 м. Талломы прикрепленной формы грациллярии в этом районе встречались только на этом плато в биотопе гальки и ракушечника. Плотность талломов грациллярии в биотопе галечников в районе мыса Большой Утриш за весь период исследования с 1985 по 1989 гг. варьировала от нуля до 10-15 экз./м2. Биомасса грациллярии на обоих полигонах за весь период наблюдений не превышала 50 г/м2. В ходе наблюдений была отмечена существенная многолетняя динамика зарослей грацилярии на одних и тех же участках акватории. Так, например, в районе мыса Большой Утриш заросли грациллярии стабильно отмечались в период с 1985 по 1989 гг., но ни разу не были отмечены в том же месте в период с 2005 по 2008 гг. Наиболее подробные исследования многолетней динамики зарослей грациллярии проводилось на акватории бухты Судак-Лиман Судакского залива Черного моря. Существенные изменения в площади зарослей и плотности талломов грациллярии отмечались после сильных штормов, при которых происходило перемещение рыхлых грунтов и, следовательно, изменение структуры вмещающего биотопа. Было отмечено, что нанос песка на прибрежные галечники происходит после штормов силой более 5 баллов южного и юго-восточного направления, а размыв галечников – после штормов силой более 5 баллов юго-западного направления. Толщина наноса и, соответственно, смыва песка при этих процессах может достигать 10-15 см. При этом перемещение более крупных фракций грунта (гальки и мелких валунов) практически не происходит. Аналогичные величины переноса песка в открытом море были получены при наблюдениях за положением стальных свай, замытых в грунт на 3 м и бетонных якорей высотой 60 см (весом до 800 кг), которые использовались на сопредельных акваториях в качестве якорной системы гидробиотехнических сооружений для промышленного культивирования мидий. В частности, было показано, что на глубине 18-25 м изменения положения уровня песка относительно замытой в грунт сваи под действием штормов может меняться на 30-40 см. Отмечено, что процесс перемещения песчаных наносов не приурочен к какому-то конкретному сезону года, хотя и тяготеет к зимнему периоду повышенной штормовой активности. Отмечено также, что после прекращения шторма не происходит автоматического восстановления структуры грунта под действием придонных течений. Длительность «песчаной» или «галечной» стадии наблюдаемого участка дна могла варьировать от несколько месяцев до несколько лет, в зависимости от повторяемости штормов с соответственных направлений. Отмечено, что после штормовых наносов песка в структуре биоценоза галечников происходили резкие изменения. Доминирование мелкозернистого песка в гранулометрическом составе грунта приводило к практически полной гибели поселений модиолы, митиллястера и теребеллид и исчезновению зарослей грациллярии. Если «песчаная» стадия длилась более полугода, то в зообентосе начинали доминировать двустворчатые моллюски Chamelia gallina, приуроченные к мелкому песку, а среди макрофитов – Polysiphonia elongata, приуроченные к раковинам хамелеи или мелкой гальке. В зависимости от периодичности, направления и силы штормов и, соответственно, величины песчаного наноса, в течение всего периода исследования наблюдались и смешанные сообщества, состоящие из видов приуроченных к биотопу галечников и к биотопу рыхлого песка. Обсуждение и выводы Таким образом, за весь период исследования структура и биологические показатели зарослей грациллярии на обследованных участках побережья соответствовали известным средним значениям для Черного моря (Миронова, 2010). Однако, под воздействием штормового переноса песка, может происходить кардинальная перестройка структуры биотопа и, соответственно, изменение структуры его биоценоза, включая, в частности, грациллярию, заросли которой могут исчезать на период от нескольких месяцев до нескольких лет. Дополнительным субстратом для роста прикрепленной грациллярии может быть рапана, обитающая в биоценозе галечников. Эти моллюски в неблагоприятные периоды года могут на несколько месяцев впадать в спячку, зарываясь в грунт на несколько сантиметров (Переладов, 2012, в печати). Дополнительным фактором для интенсивного роста грациллярии на раковинах живых рапан могут быть продукты метаболизма моллюска, содержащие биогенные элементы.
Список литературы 1. Блинова Е.И., Пронина О.А., Штрик В.А. Методические рекомендации по учету запасов промысловых морских водорослей прибрежной зоны // Методы ландшафтных исследований и оценки запасов донных беспозвоночных и водорослей морской прибрежной зоны. Выпуск 3. Изучение экосистем рыбохозяйственных водоемов, сбор и обработка данных о водных биологических ресурсах, техника и технология их добычи и переработки. М.: Изд-во ВНИРО, 2005. С. 80-127. 2. Блинова Е.И. Водоросли-макрофиты и травы морей европейской части России. М.: ВНИРО, 2007. 114 стр. 3. Вилкова О.Ю. Проблемы изучения и освоения промысловых водорослей морей России // Морские прибрежные экосистемы. Водоросли, беспозвоночные и продукты их переработки: тезисы докл. Третьей международн. научно-практ. конф. - Владивосток: ТИНРО-Центр, 2008. - С. 401-402. 4. Вилкова О.Ю., Подкорытова А.В., Котенев Б.Н. О возможности адаптации биотехнологии культивирования агарофитов в условиях Черного моря // Рыбные ресурсы. 2008. №2. С. 48-51 5. Калугина А.А., Миронова Н.В. Внутривидовая структура Gracillaria verrucosa (Huds.) Papenf в Черном море // Новости систематики низших растений. 1985. Т. 22. Л.: Наука. С. 54-59 6. Миронова Н.В. Морфо-биологическая характеристика и распостранение Gracillsria verrucosa (Huds.) Papenf. В Черном море // Экология моря. 2002. Вып.50. С.48-52. 7. Миронова Н.В. Экология, фитоценология и культивирование черноморской водоросли рода Gracillaria: Автореферат дис. … канд.биол.наук - Севастополь, ИнБЮМ, 2005. - 21 стр. 8. Миронова Н.В. Эколого-фитоценотические особенности произрастания видов рода грациллярия (обзор) // Экология моря. 2010. Спец. Вып. 81. С. 59-65. 9. Переладов М.В. Современное состояние популяции и особенности биологии рапаны (Rapana venosa) в Черном море // Труды ВНИРО, 2012, в печати. опубликовано - декабрь 2012 г.
On the influence of habitat structure on the dynamics of the Black Sea Gracillaria sp. community Mikhail V. Pereladov On the base of 1981-2008 SCUBA surveys influence of waves to sublittoral sand replacement and to structure of biotopes are discussed. It was shown that changing of ground composition was the main factor which determines dynamics of red seaweed Gracillaria fields in the Black sea coastal zone. Key words: Black sea; Gracillaria; biotope; dynamics.
Об авторе Переладов Михаил Владимирович - Mikhail V. Pereladov кандидат биологических наук pereladov@vniro.ru Корреспондентский адрес: Россия, 107140, г. Москва, ул. Верхняя Красносельская, 17; телефон (499) 264-89-83.
ССЫЛКА НА СТАТЬЮ: Переладов М.В. К вопросу о влиянии структуры биотопа на динамику зарослей черноморской грациллярии // Вопросы современной альгологии. 2012. № 2 (2). URL: http://algology.ru/116

|
|||
|
| ||