|
|
|
|
V Сабининские чтения 29 ноября 2016 - 29 января 2017 Оценка жизнеспособности трех видов микроводорослей после воздействия низких температур с криопротекторами
Харчук И.А. Irina A. Kharchuk
Институт морских биологических исследований им. А.О.Ковалевского (г. Севастополь)
УДК 582.263+582.273:57.086.13
Исследована жизнеспособность двух видов зеленых микроводорослей (Tetraselmis viridis и Chlorella vulgaris f. suboblonga) и одного вида красных микроводорослей (Porphyridium purpureum) после воздействия низких температур в течение длительного времени. Проведен анализ содержания биохимических компонентов клеток в зависимости от внесенного криопротектора. Были установлены типы криопротекторов и подобраны их концентрации, позволяющие сохранять максимальное количество жизнеспособных клеток исследованных видов микроводорослей после реактивации. Ключевые слова: микроводоросли; жизнеспособность; криоконсервация; криопротекторы; Tetraselmis viridis; Porhpyridium purpureum; Chlorella vulgaris.
Проблема сохранения и воспроизводства биоразнообразия морской среды наиболее актуальна в условиях нестабильного состояния прибрежных экосистем. Одним из методов сохранения микроводорослей является перевод клеток в состояние анабиоза. Холодовой анабиоз индуцируется в клетках гипотермией и замораживанием до низких температур. В состоянии глубокого анабиоза полностью останавливаются обменные, биохимические реакции, создаются условия для длительного хранения биологической системы с последующим полным возвратом ее к исходному состоянию в условиях нормотермии. При использовании этого метода консервации довольно редки генетические изменения, а культуры микроорганизмов, сохраняемые таким способом, оказываются менее поврежденными и имеют более высокий уровень жизнеспособности, чем при высушивании и лиофилизации (Нечаева, 2004). Использование защитных веществ, в качестве протекторов, позволяет сохранить их длительное время и увеличить количество жизнеспособных клеток. Однако для каждого вида водорослей может подходить один и тот же протектор, но в разной концентрации. Цель работы - изучить жизнеспособность клеток микроводорослей разных таксономических отделов сохраняемых в морозильной камере длительное время с использованием протекторов.
Материалы и методы Объектами исследования были штаммы водорослей Tetraselmis viridis (штамм IMBR-25), Porhpyridium purpureum (штамм IMBR-70), Chlorella vulgaris f. suboblonga (штамм IMBR-19), сохраняемые в отделе биотехнологии и фиторесурсов ИМБИ РАН. Микроводоросли культивировали в накопительном режиме, при постоянном круглосуточном освещении и автоматическом перемешивании с использованием насоса для удаления избытка кислорода из среды и равномерного прогрева всего слоя питательного раствора культуры. Интенсивность света на поверхности раствора составляла 8 кЛк. Температура среды колебалась в диапазоне 25–29°С. В качестве питательной среды для T. viridis, P. purpureum использовали среду Тренкеншу, для Ch. vulgaris – среду Тамийя. Объём среды в культиваторах составлял 5 л, при высоте слоя раствора 45 см. На стационарной фазе роста культуры микроводорослей T. viridis и Ch. vulgaris, разделяли на три равные части и к двум из них добавляли глицерин. Конечная концентрация, которого составляла 10 и 20%. Клетки продолжали культивировать ещё в течение 24 ч. Культуру клеток P. purpureum делили на три равные части: к первой доле клеток добавляли 10% глицерин, как описано выше. Ко второй доле сконцентрированных клеток прибавляли природные полисахариды, которые микроводоросли синтезировали в процессе своего роста, выделенные при центрифугировании клеток. Третья доля клеток была заморожена без протекторов. Полученную пасту водорослей помещали в герметичные пластиковые боксы и ставили в морозильную камеру (-14ºС). Перед замораживанием изымали аликвоту каждой пробы водорослей и проводили биохимические исследования. Влажность в обезвоженных культурах определяли стандартным методом доведения до постоянной массы (Методы …, 1975). Пробы обрабатывали по схеме комплексного химического анализа гидробионтов (Копытов и др., 1984). Массовую долю белка в водорослях определяли по методике Лоури (Lowry, 1951), содержание пигментов – спектрофотометрическими методами на приборе СФ-2000 (Rowan, 1989). Хлорофилл (Хл) а и b экстрагировали 100%-ным ацетоном, для расчета использовали формулы (Большой практикум…, 1975). Каротиноиды (КР) оценивали в суммарной вытяжке пигментов: во влажной культуре зелёных водорослей в области 440,5 нм, красных водорослей – 663 нм (Большой практикум…, 1975). Общее содержание липидов находили спектрофотометрическим методом с фосфованилиновым реактивом (Руководство..., 2004). Определение углеводов проводили по методике А.И. Агатовой (2004) с L-триптофановым реактивом (Руководство..., 2004). Количество свободных нуклеотидов (СН), РНК и ДНК определяли спектрофотометрическим методом (Спирин, 1958). Регистрируемые показатели химического состава выражали в пересчете на сухую массу. Для определения жизнеспособности изымали часть пробы водорослей и проводили реактивацию. Срок пребывания микроводорослей в замороженном состоянии составлял для T. viridis – 127 дней, Ch. vulgaris – 219 дней, P. purpureum – 365 дней. Выявление живых и мертвых клеток микроводорослей проводили методом витального дифференциального окрашивания клеток трипановым синим (Методы.., 1975) с помощью светового микроскопа. Одновременно учитывали количество реактивируемых клеток и определяли долю жизнеспособных клеток. Под жизнеспособностью подразумевали способность микроводорослей эндогенно поглощать краситель. Критерием жизнеспособности также был рост микроводорослей на жидких питательных средах. Для количественного учёта роста микроводорослей в культуре использовали камеру Горяева.
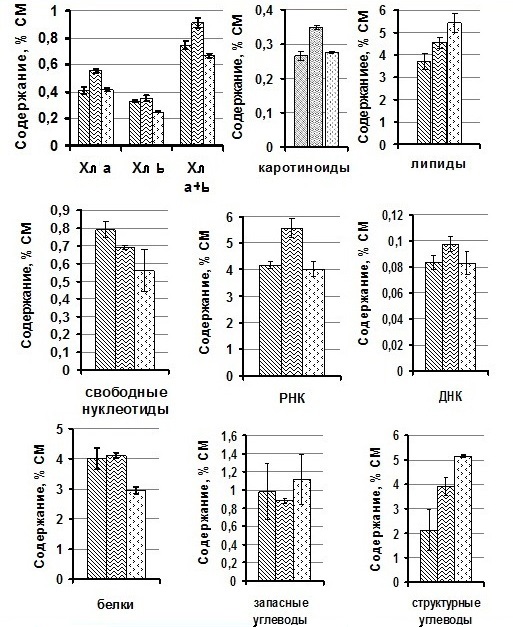
Результаты и обсуждение Анализ биохимических компонентов клеток T. viridis, показал, что в культуре с 10% глицерином статистически значимо увеличивалось содержание Хл а – на 36% и, соответственно, суммарных Хл а+b – на 23%, КР – на 34%, РНК – на 34% и ДНК – на 12% по сравнению с клетками водорослей без добавок и тех водорослей, к которым добавляли 20% глицерин (рис.1). Для проб без протектора и с 20% глицерином содержание Хл а, КР, РНК, ДНК и резервных углеводов статистически не различалось. Также выявлено, что доля липидов и структурных углеводов с увеличением концентрации глицерина, прибавляемого к клеткам T. viridis, возрастала, а содержание СН, напротив, снижалось.
Рис. 1. Содержание биохимических компонентов в клетках Tetraselmis viridis в зависимости от используемого протектора.
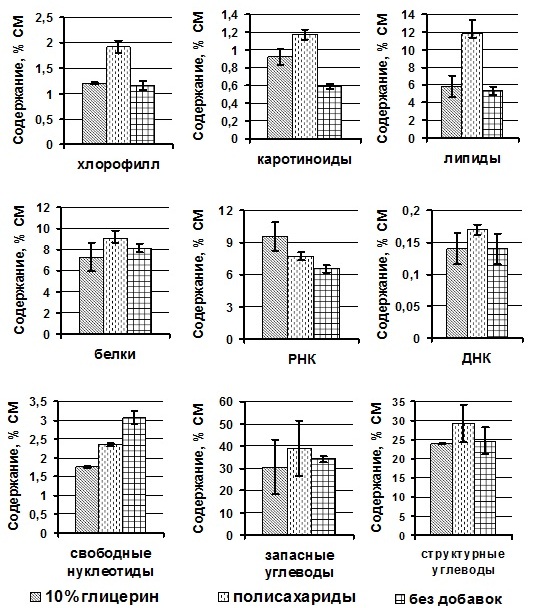
В клетках P. purpureum, сохраняемых с предварительно внесённым 10% глицерином, при отрицательной температуре (-14ºС) зарегистрировано статистически значимое повышение содержания КР – на 56%, липидов – на 10% и РКН – на 46%, по сравнению с содержанием в клетках без протекторов. Значения остальных показателей биохимического состава у сравниваемых образцов статистически не отличались (рис. 2). В клетках, замороженных с природными полисахаридами, синтезируемыми клетками P. purpureum в процессе жизнедеятельности, отмечено увеличение содержание Хл а – на 65%, КР – на 98%, липидов – на 120%, РНК – на 46% по сравнению с клетками без протекторов. При исследовании биохимических компонентов в культурах, сохраняемых с 10% глицерином и полисахаридами, зарегистрировано статистически значимое возрастание доли Хл а – на 60%, КР – на 27%, липидов – на 100% и структурных углеводов – на 22% по сравнению с клетками, заложенными на хранение с дополнительно внесёнными полисахаридами. Характерной особенностью было то, что во всех культурах, независимо от добавления протектора, значения ДНК и белков были статистически одинаковы.
Рис. 2. Содержание биохимических компонентов в клетках Porphyridium purpureum в зависимости от используемого протектора.
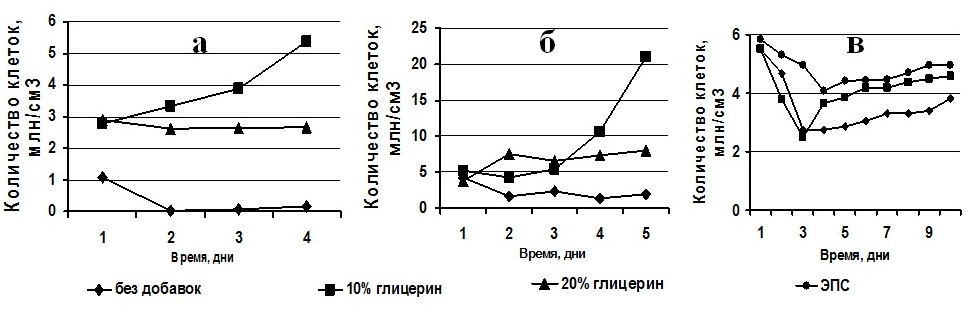
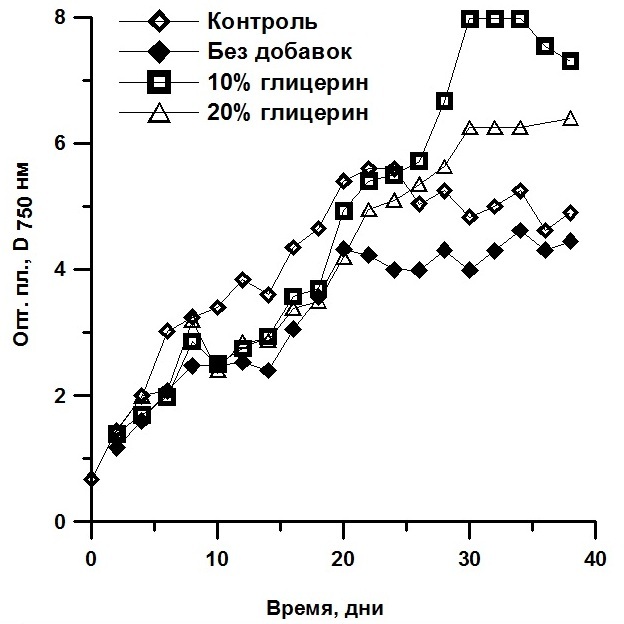
Клетки T. viridis, Ch. vulgaris и P. purpureum были реактивированы после 4, 7 и 12 месяцев хранения в морозильной камере соответственно. Динамика жизнеспособных клеток в процессе реактивации представлена на рис. 3. Реактивация T. viridis через 127 суток хранения в морозильной камере показала, что количество жизнеспособных клеток в пробах, сохраняемых с глицерином, было в 2,5 раза выше, чем в пробе без добавления протектора (рис. 3а). На второй день количество клеток в исследуемых образцах, сохраняемых с 20% глицерином и без него, снизилось на 9,32% и 97% соответственно. В то время как в образце, замороженном с 10% глицерином, количество клеток увеличилось на 16,6%. На третий и последующие дни реактивации во всех пробах отмечено увеличение количества жизнеспособных клеток за счёт их деления.
Рис. 3. Динамика жизнеспособных клеток в процессе реактивации культур: а) Tetraselmis viridis, б) Chlorella vulgaris, в) Porphyridium purpureum.
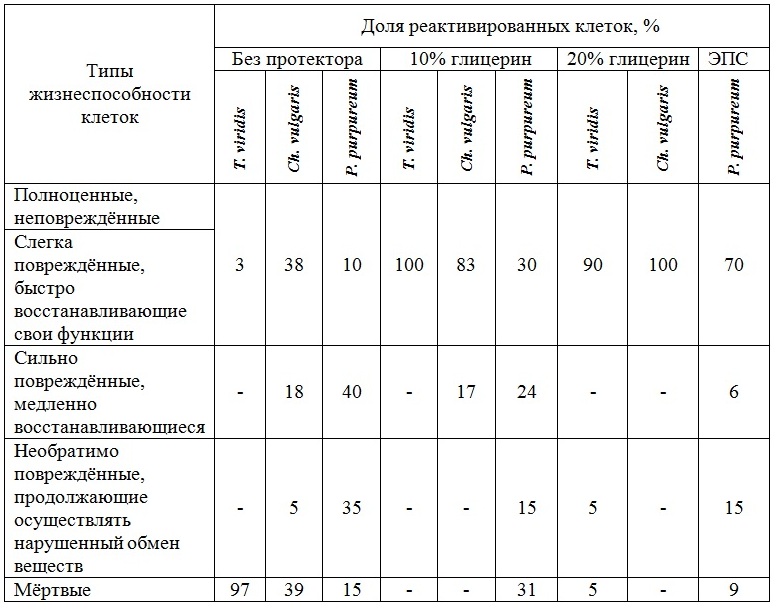
Во время реактивации клеток Ch. vulgaris, сохраняемых при температуре -14°С в течение 219 суток, зарегистрировано, что на второй день количество клеток в исследуемых образцах, сохраняемых с 10% глицерином и без него, снизилось на 17% и 62% соответственно (рис. 3б). В то время как в образце, сохраняемом с 20% глицерином, количество клеток увеличилось на 47%. На третий день реактивации в образцах, предварительно сохраненных с добавлением 10% и 20% глицерина, число жизнеспособных клеток увеличилось на 25% и 18% соответственно, а в культуре водорослей, сохраняемой без криопротектора, число жизнеспособных клеток, относительно предыдущего дня, увеличилось на 46%. Во всех пробах отмечено деление клеток. Клетки P. purpureum были реактивированы через 365 суток пребывания в морозильной камере. В процессе их оживления (на 3–4 день) отмечено снижение жизнеспособных клеток в 2 раза в пробах, сохраняемых без добавок и с 10% глицерином и в 1,4 раза в пробе с ЭПС (рис. 3в). На 6–7 сутки реактивации количество клеток увеличилось на 20%, 69% и 9% соответственно. Анализ полученных данных согласно классификации клеток по жизнеспособности (Мейсель, 1961) показал, что в реактивированных пробах, замороженных как без протекторов, так и с протекторами, присутствуют клетки 5 типов (табл. 1), при этом разделение на 1 и 2 тип условно и для каждого вида водорослей специфично. Такое распределение клеток связано с тем, что при низких температурах идет переход мембранообразующих липидов из жидкокристаллического состояния в гелеобразное, это приводит к образованию весьма ригидной мембранной структуры, пластичность которой резко отличается от состояния мембран, находящихся при физиологических температурах. При замораживании, из-за растущих внеклеточных кристаллов, идет сильное сдавливание и деформация клеток, все это в конечном итоге приводит к появлению многих разрывов в бислойной структуре клеточной мембраны и «выдавливанию» белков из мембраны, соответственно к гибели клеток (Скорняков, 1980). Известно, что избежать образования льда можно снижением точки замерзания раствора путем накопления электролитов или спиртов типа глицерина и углеводов – сахаров. Глицерин – протектор проникающего действия, он препятствует формированию кристаллов за счёт образования водородных связей с молекулами воды. Он стабилизирует аминокислоты и способствует сохранению генетического аппарата (Назаренко, 2008). T. viridis и Ch. vulgaris – зелёные микроводоросли, которые в процессе жизнедеятельности могут синтезировать глицерин при неблагоприятных для них условиях (Balnokin, 2004), а также ассимилировать его из культуральной среды, переходя к миксотрофному типу питания (Liang, 2009, Kong et al., 2013, Жондарева, 2016), вероятно из-за этой способности происходит увеличение количества липидов в клетках T. viridis при добавлении к ним глицерина. Чем больше количество добавляемого глицерина, тем выше концентрация липидов в клетках, это приводит к повышению протекторных свойств клеток. Меньше повреждаются клеточные структуры при замораживании, соответственно увеличивается количество структурных углеводов, РНК и ДНК, а доля свободных нуклеотидов снижается. Экзополисахариды (ЭПС), синтезируемые и выделяемые клетками P. purpureum в процессе их жизнедеятельности, являются естественными криопротекторами, их ещё называют антифризами, т.к. в их состав входят ксилоза, галактоза и глюкоза, эти компоненты стабилизируют клеточные мембраны и содержимое клеток (Назаренко, 2008). Считается, что действие антифризов основано на наличии в их молекулах большого числа полярных группировок. Таким образом, возможно с помощью антифризов осуществляется блокада роста кристаллов льда. Вероятно поэтому ЭПС – наиболее эффективные консервирующие субстанции для P. purpureum. По результатам эксперимента определено, что доля клеток, сохранивших репродукционные свойства, у T. viridis в пробе без добавок составляла 3%, в пробе с 10% глицерином – 100%, с 20% глицерином – 90%; у Ch. vulgaris в пробе без добавок составляла – 38%, в пробе с 10% глицерином – 83%, с 20% глицерином – 100%; у P. purpureum в пробе без добавок составляла – 10%, в пробе с 10% глицерином – 30%, с ЭПС – 70%.
Таблица 1. Классификация по жизнеспособности реактивированных клеток микроводорослей, сохраняемых с протекторами Table 1. Types of reactivated microalgae cells (keept with protectors) according to their viability
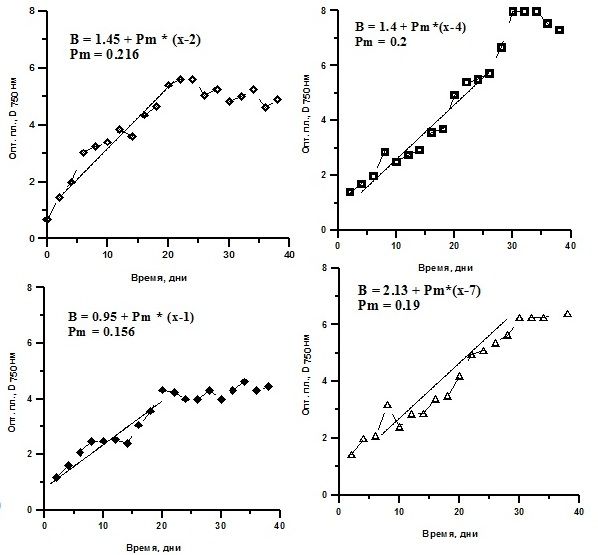
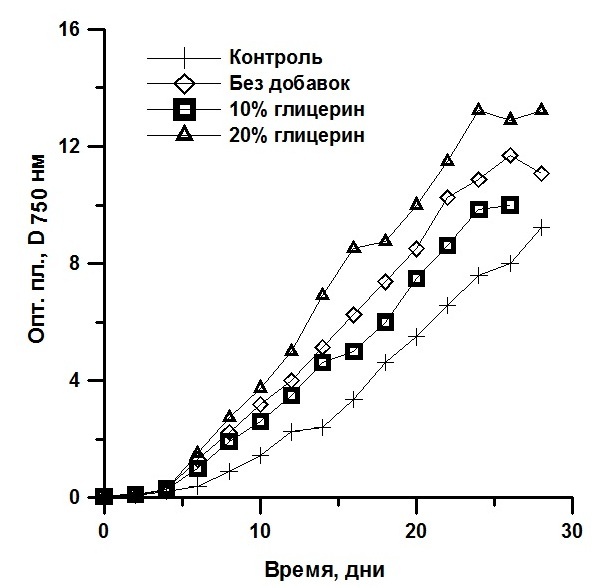
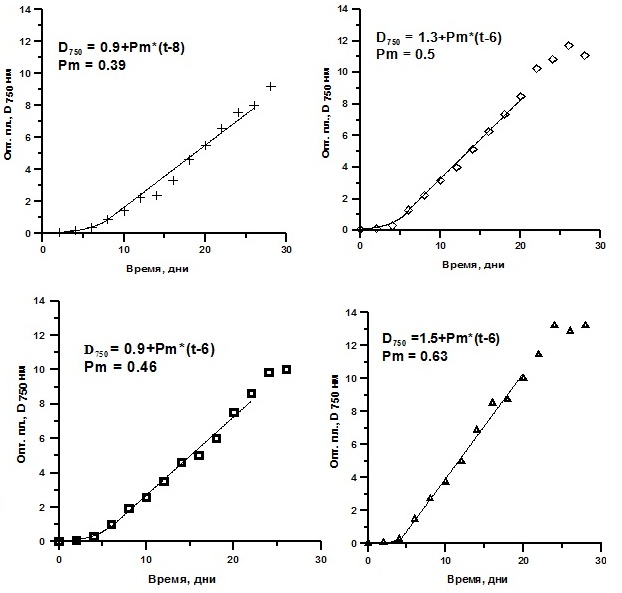
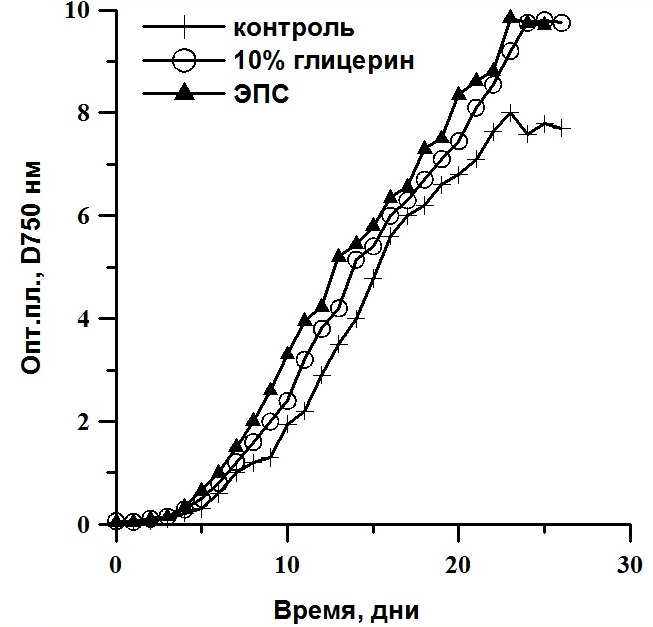
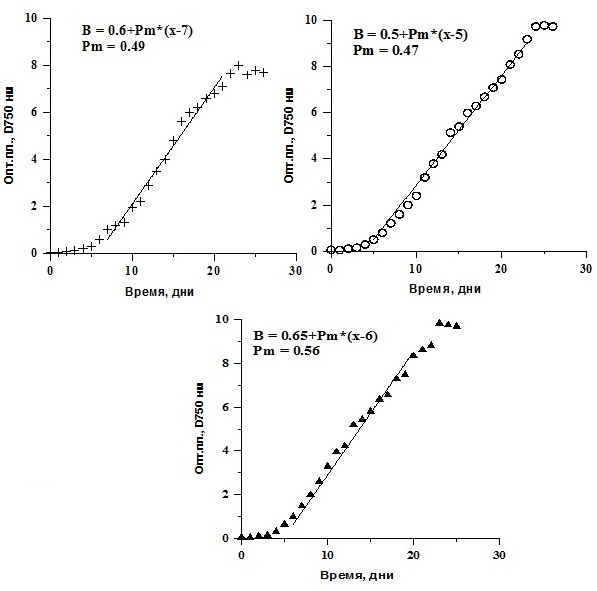
Реактивированные клетки были выведены в культуру. Далее их выращивали накопительным способом. Ростовые характеристики реактивированных микроводорослей T. viridis, Ch. vulgaris и P. purpureum были практически идентичны контролю (культуры, не подвергавшиеся замораживанию) и не зависели от протектора и его концентрации (рис. 4, 5, 6). Исследование динамики накопления биомассы реактивированных культур показало, что кривые роста идентичны. Максимальные значения средней продуктивности на линейной стадии роста у T. viridis были выше в культурах подлежащих замораживанию с глицерином, у Ch. vulgaris – с глицерином в концентрации 20%, у P. purpureum – с ЭПС.
Рис. 4. Динамика биомассы накопительных культур Tetraselmis viridis после реактивации. D750 – динамика плотности (ед.опт.пл D750*сут), Рm – средняя продуктивность на линейной фазе роста (г СМ/л*сут).
Рис. 5. Динамика биомассы накопительных культур Chlorella vulgaris после реактивации. D750 – динамика плотности (ед.опт.пл D750*сут), Рm – средняя продуктивность на линейной фазе роста (г СМ/л*сут).
Рис. 6. Динамика биомассы накопительных культур Porhpyridium purpureum после реактивации. D750 – динамика плотности (ед.опт.пл D750*сут), Рm – средняя продуктивность на линейной фазе роста (г СМ/л*сут).
Были проведены эксперименты, связанные с изучением жизнеспособности двух видов зеленых микроводорослей (Tetraselmis viridis и Chlorella vulgaris f. suboblonga) и одного вида красных микроводорослей (Porphyridium purpureum) после воздействия низких температур (-14°С) в течение длительного времени (от 127 до 365 суток). Показано, что глицерин позволяет сохранить высокое количество жизнеспособных клеток зелёных микроводорослей после реактивации, в частности, для сохранения 100% жизнеспособных клеток для T. viridis необходимо использовать 10% глицерин, для Ch. vulgaris – 20% глицерин. Для P. purpureum лучшим криопротектором являются экзополисахариды, синтезируемые самой водорослью, использование которых при замораживании и хранении предохраняет от разрушения 70% клеток.
Работа выполнена в рамках госзадания № 1001-2014-0017 «Разработка научных основ решения гидробиологических и биотехнологических проблем интегрированного управления прибрежными зонами».
Список литературы 1. Большой практикум по физиологии растений / Б.А. Рубин (ред.) – М.: Высш. Шк., 1975. – 439 с. 2. Жондарева Я.Д., Тренкеншу Р.П. Фотоавтотрофный и миксотрофный рост Tetraselmis viridis в накопительной культуре // Актуальные вопросы биологической физики и химии. 2016. №1–2. С. 106–109. 3. Копытов Ю.П., Дивавин И.А., Цымбал И.М. Схема комплексного биохимического анализа гидробионтов // Материалы конф. «Рациональное использование ресурсов моря – важный вклад в реализацию продовольственной программы» (Севастополь, 10–11 дек. 1984). – Севастополь, 1984. – Ч.2. – C. 227––231. – Деп. в ВИНИТИ 16.04.85, № 2556 – 85. 4. Мейсель М.Н., Медведева Г.А., Алексеева В.М. О выявлении живых, повреждённых и мертвых микроорганизмов // Микробиология. 1961. Т.30, №5. С. 855–862. 5. Методы физиолого-биохимического исследования водорослей в гидробиологической практике / А.В. Топачевский (отв. ред.). – Киев: Наук. думка, 1975. – 247 с. 6. Назаренко Л.В. Некоторые особенности криосохранения биологических объектов // Вестник Московского городского педагогического университета. Серия: Естественные науки. 2008. №1. С. 53–59. 7. Нечаева О.В. Поиск и использование синтетических соединений гетероциклического ряда для сохранения стабильности популяционного состава коллекционных культур микроорганизмов: дис. ... канд. биол. наук. – Саратов, 2004. – 143 с. 8. Руководство по современным биохимическим методам исследования водных экосистем, перспективных для промысла и марикультуры / А.И. Агатова (ред.). – М.: Изд-во ВНИРО, 2004. – 123 с. 9. Скорняков Б.А., Вишневский В.И. Особенности механизма крио-протекции глицерином // Криобиология и криомедицина. 1980. №6. С. 59–62. 10. Спирин А.С. Спектрофотометрическое определение суммарного количества нуклеиновых кислот // Биохимия. 1958. Т.23, №5. С. 656–662. 11. Balnokin Y.V. The Na+-translocating ATPase in the plasma membrane of the marine microalga Tetraselmis viridis catalyzes Na+/H+ exchange / L.G. Popova, L.Y. Pagis, I.M. Andreev // Planta. 2004. Vol.219, №2. P. 332. 12. Kong W.B., Yang H., Cao Y.T., Song H., Hua S.F., Xia C.G. Effect of glycerol and glucose on the enhancement of biomass, lipid and soluble carbohydrate production by Chlorella vulgaris in mixotrophic Culture // Food technol. Biotechnol. 2013. 51, 1. Р. 62–69. 13. Liang Y., Sarkany N., Cui Y. Biomass and lipid productivities of Chlorella vulgaris under autotrophic, heterotrophic and mixotrophic growth conditions // Biotechnol Lett. 2009. 31, №7. Р. 1043–1049. 14. Lowry O.H., Rosebrough N.J., Farr A.L., Randall R.J. Protein measurement with folin phenol reagent // Journ. Biol. Chem. 1951. 193, №1. P. 265–275. 15. Rowan K.S. Photosynthetic Pigments of Algae – Cambridge: Cambridge Univ. Press, 1989. – 334 p. Статья поступила в редакцию 20.12.2016
Assessment of viability of three species of microalgae after low-temperature exploration with cryoprotectors Irina A. Kharchuk The viability of two species of green microalgae (Tetraselmis viridis and Chlorella vulgaris f. suboblonga) and a species of red microalga (Porphyridium purpureum) after low-temperature exploration for a long time was investigated. The analysis of the content of biochemical components of cells depending on added cryoprotectant was made. The types of cryoprotectants were determined and their concentrations were chose to allow preservation of a maximum number of viable cells of three aforementioned microalgae after their reactivation. Keywords: microalgae; viability; cryopreservation; cryoprotectants; Tetraselmis viridis; Porhpyridium purpureum; Chlorella vulgaris.
Об авторе Харчук Ирина Алексеевна - Kharchuk Irina A. кандидат биологических наук seaferm@yandex.ru Корреспондентский адрес: Россия, 299011, Севастополь, пр. Нахимова, 2, ФГБУН ИМБИ; тел. (869)-2550795.
ССЫЛКА НА СТАТЬЮ: Харчук И.А. Оценка жизнеспособности трех видов микроводорослей после воздействия низких температур с криопротекторами // Вопросы современной альгологии. 2017. № 1 (13). URL: http://algology.ru/1133 Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно.
При перепечатке ссылка на сайт обязательна
К разделу ОБЗОРЫ, СТАТЬИ И КРАТКИЕ СООБЩЕНИЯ

|
|||
|
| ||