|
|
|
|
по Материалам Международной конференции «Экологическая физиология водных фототрофов: распространение, запасы, химический состав и использование» V Сабининские чтения 29 ноября 2016 - 29 января 2017 Видовой состав, экологическая структура и количественная характеристика макроводорослей бухты Голландия (Черное море)
Евстигнеева И.К., Танковская И.Н. Irina К. Evstigneeva, Irina N. Tankovskaya
Институт морских биологических исследований им. А.О.Ковалевского (г. Севастополь)
УДК 582.275.54+581.55
Впервые описаны видовой состав и экологическая структура макрофитобентоса и макрофитоперифитона черноморской бухты Голландия в месте размещения полигона Центра морских исследований и технологий СевГУ. Выделены эколого-таксономические лидеры и главные продуценты. Выявлены основные тенденции пространственных изменений макрофитобентоса и макрофитоперифитона бухты. Ключевые слова: Черное море; бухта Голландия; макрофитобентос; макрофитоперифитон; встречаемость; эколого-таксономическая структура; фитомасса; изменчивость.
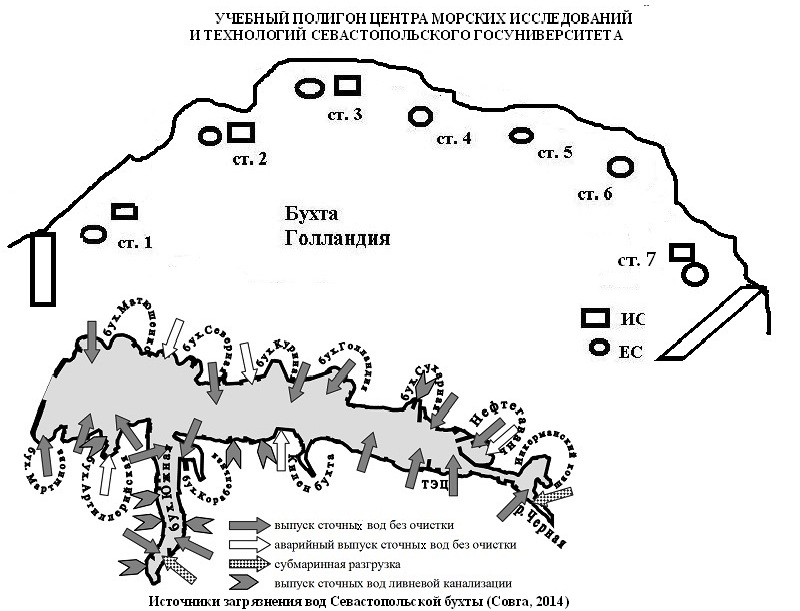
Севастопольская бухта относится к числу морских акваторий, подверженных антропогенному воздействию. Она является градообразующим элементом геосферы, служит базой черноморского флота РФ со своей промышленно-производственной и хозяйственной инфраструктурой, принадлежит к зоне активного судоходства и осуществления гидротехнических работ, что приводит к нарушению существующего равновесия. Ежедневно в бухту сбрасывается до 10–15 тыс. м3 неочищенных или условно-чистых вод с широким спектром загрязняющих веществ в концентрациях, значительно превышающих допустимые нормы (Совга и др., 2014). Структурным элементом бухты Севастопольская является бухта Голландия, часть прибрежья которой занята учебным полигоном Центра морских исследований и технологий Севастопольского госуниверситета (рис. 1). Вблизи находится стоянка кораблей, причал городских катеров, действует выпуск сточной воды без очистки и функционирует стихийный городской пляж. Такая нагрузка не может не сказаться на условиях жизни гидробионтов, их видовом разнообразии и структурно-функциональных характеристиках образуемых ими сообществ.
Рис. 1. Карта – схема района исследований (ИС – искусственный субстрат, ЕС – естественный субстрат)
Важной составляющей морских донных ценозов является макрофитобиота. Ее первое исследование в бухте Голландия относится к середине прошлого столетия (1967 и 1977 гг.) и касается только фитосапробного состава макроводорослей. Известно, что к 1977 году, в связи со строительством портовых сооружений, в бухте Голландия растительность исчезает (Калугина-Гутник, 1982). В начале текущего столетия нами были продолжены и расширены исследования, касающиеся эколого-таксономического состава и фитомассы морской растительности этого района (Евстигнеева, Танковская, 2004; Евстигнеева, 2006). В 2016 году гидроботанические изыскания возобновляются в связи с организацией здесь Центра морских исследований и технологий со своим учебным полигоном. Цель работы – исследование эколого-таксономического состава, структуры, продукционных возможностей и пространственной динамики макрофитобентоса (МФБ) и макрофитоперифитона (МФП) твердых субстратов бухты Голландия.
Материалы и методы Объектом исследования стали макроводоросли искусственного (ИС) и естественного субстратов (ЕС) бухты в районе размещения учебного полигона Центра морских исследований и технологий. Полигон по отношению к входу в бухту частично охватывает вершину бухты, полностью – правый берег водоема и ограничен причальными конструкциями. В августе 2016 г. пробы отбирали вручную на семи станциях (ст.) методом учетных площадок (25х25 см) (Калугина-Гутник, 1969) в зоне верхней сублиторали на одной и той же глубине (до 0,2 м). Ст. 1 и 2 приурочены к вершине бухты, ст. 3 и 4 с двух сторон примыкают к местному пляжу, ст. 5 и 6 располагаются вдоль правого берега, ст. 7 находится за причалом городских катеров вблизи выхода из бухты. МФП развивается на бетонных плитах, укрепляющих берег на ст. 1, 2, 3 и 7. При камеральной обработке проб определяли видовой состав макроводорослей с учетом последних номенклатурных изменений (Зинова, 1967; Guiry, Guiry, 2016). Для описания видовой структуры рассчитывали коэффициенты встречаемости (R, %) и флористического сходства по Жаккару (Kj, %) (Розенберг, 2005), а для экологической – применяли шкалу А.А. Калугиной-Гутник (Калугина-Гутник, 1975). Исходя из значений коэффициента R, виды распределяли на группы постоянства. По индивидуальной фитомассе выявляли видовой состав доминантов и содоминантов, рассчитывали индекс их доминирования (Dt, %), представляющий отношение числа проб, где вид доминировал по фитомассе, к числу проб, где он встречался. Для коррекции списка доминирующих видов применяли шкалу Е.Л. Любарского (Розенберг, 2005). Для описания изменчивости характеристик МФБ и МФП определяли лимиты и размах их вариации, а по величине коэффициента вариации (Сv, %) – тип изменчивости биологических признаков в соответствии со шкалой Г.Н. Зайцева (Зайцев, 1990). Для оценки качества морских вод был применен индекс сапробности (Калугина-Гутник, 1989).
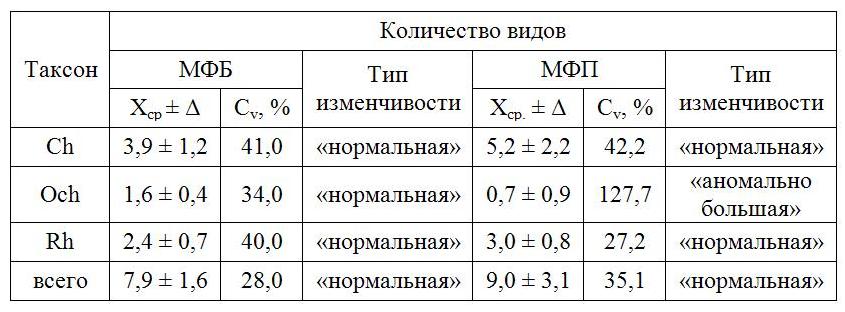
Результаты и обсуждение Общая таксономическая характеристика МФБ и МФП акватории полигона Визуальное обследования морской части полигона показало, что макроводоросли заселяют как прибрежные камни, небольшие валуны, так и бетонные плиты, укрепляющие берег. Проективное покрытие твердых субстратов водорослями на первой станции (для искусственного субстрата еще и на ст. 2) составляет 100%, на остальных – варьирует от 70 до 90%. В конце лета происходит смена поколений у такой ценозообразующей черноморской водоросли, как Cystoseira barbata C. Ag. Ее молодые проростки длиной 2–3 см размещаются на границе суши и моря, в зоне заплеска. Для макрофитов бухты на обоих субстратах характерен выраженный эпифитизм. Причем эпифитные формы водорослей вегетируют даже на проростках. В ходе исследований в составе донных сообществ бухты Голландия обнаружены 15 видов макроводорослей, относящихся к 9 родам, 7 семействам и 6 порядкам классов Ulvophyceae отдела Chlorophyta (Ch), Phaeophyceae отдела Ochrophyta (Осh) и Florideophyceae отдела Rhodophyta (Rh). Из них в состав МФБ и МФП входят 15 и 14 видов, соответственно (табл. 1). Среди отделов по таксономическому разнообразию лидирует Ch (53% общего числа идентифицированных видов), что является характерной чертой растительности твердых субстратов этого водоема, тогда как в большинстве других районов Черного моря такое положение, как правило, занимают Rh. Количество видов Rh в 2,5 раза больше, чем Осh и в 1,6 раза меньше, чем Ch. Такое распределение видов между отделами можно рассматривать как адаптационную реакцию водорослей и их сообществ на условия обитания в бухте. Основу таксономического разнообразия донной альгофлоры бухты образуют такие роды, как Cladophora (3 вида) и Ulva (4 вида) из Ch. Среди Rh род Ceramium представлен двумя видами, остальные – одним (табл. 1). Доля моновидовых родов в общем списке составляет 40%. Полученные данные свидетельствуют о том, что таксономическая структура Осh среди отделов наиболее упрощенная, Ch занимает ведущую позицию по количеству видов, а Rh – родов. Встречаемость макроводорослей в прибрежье полигона С учетом величины коэффициента R макроводоросли ЕС бухты распределены на такие группы, как постоянная (R >50%), добавочная (R = 25-50%) и случайная (R< 25%) с преобладанием первой (46% видов). Остальные группы одновременно занимают второе место (по 27%). Виды ИС поровну распределяются между постоянной и добавочной группами. Сравнительный анализ структурно-функциональных характеристик МФБ и МФП и их пространственная динамика Флористическая структура МФБ и МФП бухты во многом соответствует друг другу. Это, прежде всего, касается уровня видового разнообразия и количественного распределения видов по отделам. Независимо от топической приуроченности видовая пропорция отделов выглядит одинаково – 4 Ch : 1 Осh : 2 Rh. Основа надвидового разнообразия каждого сообщества сформирована одними и теми же таксонами. Значения коэффициента Kj для всего видового состава фитоценозов (93%), а также для Ch (100%), Осh (100%) и Rh (80%) в отдельности свидетельствуют о высокой качественной однородности МФБ и МФП бухты. На разных вдольбереговых станциях наибольшее сходство характерно видовому составу Осh МФБ (71%), наименьшее – его Ch (49%). Такая же тенденция характерна и для МФП. Минимум качественных совпадений проявляет видовой состав МФБ на ст. 3 при его сравнении с таковым на ст. 6 и 7 (39 и 36%). Максимум видового сходства МФБ чаще приходится на такие пары удаленных друг от друга станций, как ст. 1 – 4, 1 – 5, 2 – 5. Видовая структура Ch на ЕС заметно отличается на пространственно разобщенных участках и проявляет сходство на смежных (67%). Видовой состав Осh на разных участках берега одинаков полностью или наполовину. Выраженное качественное различие Rh характерно для фитоценозов на ст. 3 и 5, 3 и 6 (Kj = 33%), которое гораздо ниже на ст. 1 и 2, 1 и 4, 2 и 4. На ИС в районе ст. 3 и 7 видовой состав МФП совпадает лишь на треть (Kj = 36%), а на ст. 1 и 2, 2 и 7 такое сходство распространяется на 72% видов. Треть общих видов Ch можно было обнаружить при сопоставлении водорослей ст. 1 и 3, 2 и 3, еще больше – на ст. 1 и 2, 1 и 7 (71 и 86%). У Осh в составе МФП пики сходства и различия примерно те же, что и в МФБ. Видовая структура Rh проявляет особенное своеобразие при сравнении тех же станций, где ранее было зафиксировано высокое качественное совпадение у Ch МФП (ст. 1 и 7) и очень низкое – у всего этого сообщества (ст. 3 и 7). Процедура сопоставления видового разнообразия сообществ МФБ и МФП, а также слагающих их отделов, на разных станциях показала следующее. На ст. 1 у вершины бухты видам Осh характерно максимально высокое сходство, тогда как у Rh и сообществ в целом оно вполовину ниже, а у Ch охватывает менее трети видов. На ст. 2 видовой состав сообществ и входящих в них отделов совпадает наполовину или около того. Еще ниже подобие на ст. 3, причем среди Осh одинаковые виды отсутствуют. На ст. 7, наиболее приближенной к открытой части бухты, оба сообщества на 67% состоят из сходных видов. При этом видовой состав Rh, Ch и Осh в отдельности идентичен на 50, 71 и 100% соответственно. В целом, МФП и МФБ сильно отличаются друг от друга только на ст. 3. Располагая данными о видовом составе МФБ в бухте Голландия, а также в прилегающей к ней бухте Северная и в акваториях расположенных примерно напротив Ушаковой балки и Килен-бухты, полученными более 10 лет назад, можно сделать выводы об изменениях, произошедших за истекший срок. Воспользуемся таким инструментом, как расчетный коэффициент флористического сходства. В прошлый период значения коэффициента для перечисленных водных объектов колебались от 31 до 47% с минимумом для видового состава МФБ бухты Голландия и Ушаковой балки и максимумом – бухт Северная и Голландия. Спустя годы видовой состав в прибрежье бухты Голландия сохранился лишь наполовину (47%), что не могло не сказаться на степени аналогии видового состава МФБ вышеупомянутых участков Севастопольской бухты. Действительно, значения Kj существенно снижены (21-28%). Высокое качественное расхождение фитоценоз проявляет в обеих бухтах (снижение на 21%). Абсолютное число видов в фитоценозах и отделах подвержено пространственной изменчивости разной выраженности. Общее число видов МФБ меняется от 5 (ст. 3) до 11 (ст. 7), составляя в среднем 7,9±1,6. Величина коэффициента Cv (28%) свидетельствует о том, что пространственные изменения видового состава МФБ являются «верхненормальными». Больше всего видов обитает на ЕС правого берега (ст. 5-7), меньше – на станциях, ограничивающих пляж (ст. 3 и 4). Количественная трансформация видового состава Ch касается 3–6 видов с достижением среднего значения, равного 3,9±1,2. При этом наблюдается увеличение видового разнообразия отдела с удалением от пляжа в сторону открытой части бухты. Видовое разнообразие Осh ограничивается 1–2 видами с максимумом на станциях, расположенных ближе к вершине бухты. Изменчивость числа видов Осh, подобно Rh, является «нормальной». Число видов Rh минимально на ст. 3 и максимально на ст. 7. Общая направленность количественных изменений видового состава данного отдела соответствует таковым у Ch (табл. 2).
Таблица 2. Параметры изменчивости видового состава МФБ и МФП в бухте Table 2. Parameters of variability of macrophytobenthos species composition and macrophytoperiphyton in the bay
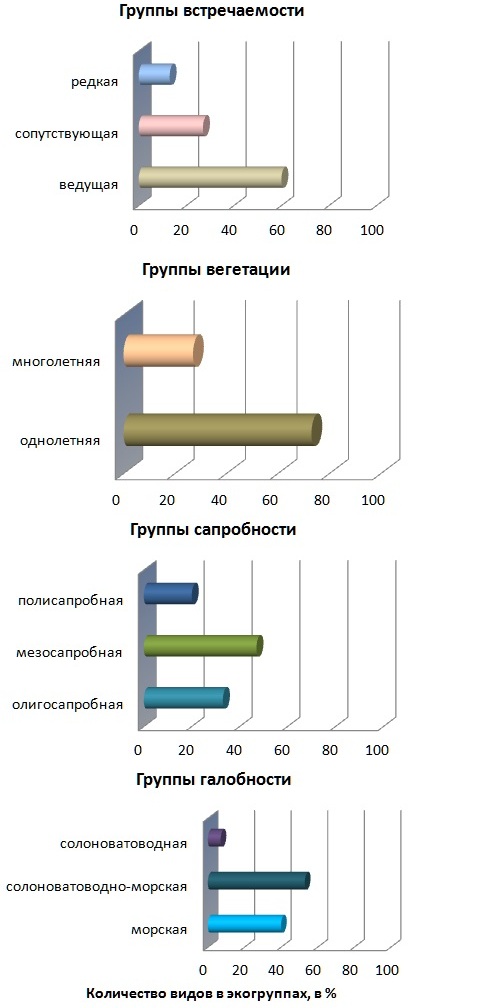
В конечном итоге, все преобразования числа видов в МФБ и отделах носят «верхненормальный» характер. Минимум видового разнообразия всего фитоценоза, Rh и Ch приходится на ст. 3 (у Осh на смежной ст. 4), а максимум – на ст. 6 и 7. Пиковое значение числа видов Осh зарегистрировано сразу на нескольких участках полигона (ст. 1–3 и 5). Лимиты пространственной изменчивости видового состава отделов и фитоценоза, формирующегося на ИС, мало отличимы от таковых на ЕС. В то же время среднее число видов всего МФП, а также его Rh и Ch на 20–33% выше, чем в МФБ. Кроме того, среднее число фитоперифитонных видов Осh на четверть меньше фитобентосных. Установлено, что максимум видового разнообразия каждого из отделов и всего МФП приходится на рядом расположенные ст. 1 (у Осh) и 2 (у оставшихся отделов и всего сообщества) и, как правило, территориально не совпадает с пиковым значением у компонентов МФБ. Минимумы видового разнообразия Ch и всего МФП зафиксированы, соответственно, на ст. 2 и 3. У Осh они приходятся сразу на несколько станций, среди которых и ст. 7, где такой уровень разнообразия характерен для этого отдела и на ЕС. На этой станции обитает наименьшее число видов Rh, примерно столько же видов и на ст. 3, где также мало и бентосных видов отдела. Экологическая структура донной растительности прибрежья бухты Голландия представлена одиннадцатью группами из тринадцати, известных для бентали Черного моря (рис. 2). На ЕС среди галобных групп наибольшее развитие получают виды солоноватоводно-морской, ведущей, однолетней и мезосапробной групп (47–73% общего числа видов). Доминирование солоноватоводно-морских и мезосапробных видов является отличительной особенностью донной фитобиоты района исследования по сравнению с большинством других участков крымского прибрежья Черного моря. Суммарная доля солоноватоводно-морской и солоноватоводной группы свидетельствует о наличии распреснения воды в прибрежье водоема (рис. 2). Выявлено, что в галобной части Ch и Rh полностью или с небольшим преимуществом над морскими видами господствуют солоноватоводно-морские растения. Первый из названных отделов представлен тремя галобными группами, второй – двумя (отсутствует солоноватоводная группа). Представители Осh относятся исключительно к морским видам.
Рис. 2. Доля видов (% их общего числа) разных экогрупп в донной растительности бухты Голландия
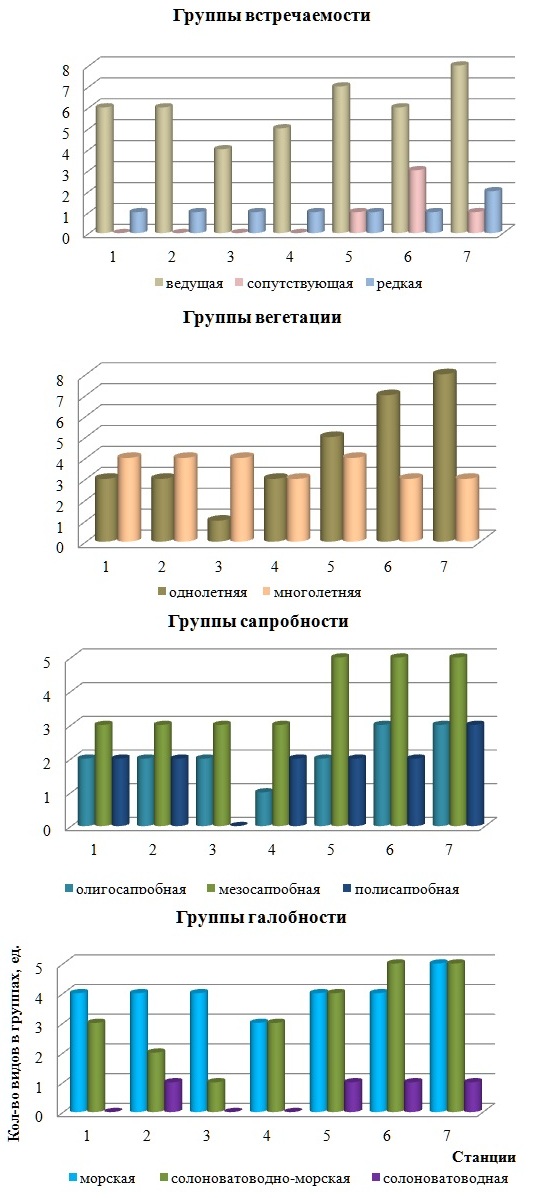
Абсолютное число видов морской группы колеблется в узких пределах (3-5 видов) с минимумом на ст. 4 и максимумом на ст. 7, достигая в среднем 4,0±2,2 вида (рис. 3). Пространственные изменения этого параметра происходят по типу «значительных». У солоноватоводно-морской группы число видов вдоль исследованной части берега варьирует от 1 до 5 с минимумом на ст. 3 и максимумом на двух последних станциях. В отличие от названных галобных групп представители солоноватоводной группы зарегистрированы не на всех станциях, что сказалось на величине коэффициента вариации числа видов, определяющей изменчивость анализируемого показателя как «очень большая». В целом, максимум абсолютного числа видов в трех галобных группах на ЕС, за небольшим исключением, приурочен к двум последним станциям, наиболее приближенным к открытой части бухты. Количественное распределение видов водорослей МФБ по группам встречаемости в Черном море происходит следующим образом: 60% видов входят в ведущую группу, 27% – в сопутствующую и остальные – в редкую. Среди Ch превалируют ведущие виды (50%), остальные группы встречаемости представлены равной долей. Виды Осh относятся исключительно к ведущим, которые среди Rh имеют хоть и небольшое, но преимущество над сопутствующими. На ЕС бухты редкие виды отделов Осh и Rh не обнаружены. Подобно галобным группам абсолютное число видов в группах встречаемости максимально на последних двух станциях. Пространственные количественные изменения ведущих видов, как базовой группы, незначительны («нижняя» норма). Редкая группа отличается «верхненормальным» типом изменчивости числа видов, а сопутствующая – «аномально высоким», что связано с ее низкой встречаемостью вдоль берега. Виды МФБ с разной продолжительностью вегетации распределяются между многолетней и однолетней группами с заметным преобладанием последней (71%). Встречаемость каждой группы максимально высокая. Ch и Rh в основном представлены однолетниками, Осh – исключительно многолетниками. Количественные изменения в этих группах различаются: у многолетней они происходят в пределах «нижней» нормы, у однолетней они «значительные». Пиковые значения числа видов в абсолютном исчислении в группах территориально разобщены: на участке берега между ст. 3 и 7 включительно у однолетников наблюдается постепенное увеличение параметра (рис. 3). Эти же изменения у многолетников более сглаженные, со слабо выраженными пиками за пределами пляжа (ст. 1, 2, 3 и 5).
Рис. 3. Экологическая структура МФБ на разных станциях
Среди трех сапробных групп мезосапробионты доминируют не только в общем составе, среди Ch и Rh, но и на каждой станции. Изредка они и олигопробионты представлены в равной мере. Эти группы в составе МФБ встречаются на всем протяжении исследованного берега, полисапробионты отсутствуют на ст. 3. Пространственные изменения абсолютного числа видов в первых двух группах относятся к «верхненормальным», в третьей – к «значительным». В целом, с учетом величины абсолютного числа видов, среди донной растительности полигона преобладают виды ведущей, солоноватоводно-морской, однолетней и мезосапробной групп. Каждый отдел в качестве базовых элементов обязательно включают ведущие виды. Комбинации групп, доминирующих среди Ch и Rh, проявляют абсолютное сходство. Анализ пространственных изменений абсолютного числа видов показал приуроченность его пиковых значений у большинства групп примерно к одному и тому же участку полигона (правый берег, вне пляжной зоны). Все изменения показателя сводятся к шести типам с преобладанием «верхненормального» и «значительного». Если брать во внимание уровень относительного числа видов МФБ, то на всех станциях постоянными лидерами являются ведущая и мезосапробная группы. Подобное положение занимает и морская группа, но только на станциях, расположенных со стороны выхода из бухты, слева от пляжа. На других участках доля морских видов не отличается от таковой у солоноватоводно-морских. Многолетняя группа, как и морская, превалирует на первых трех станциях, на других участках берега она уступает однолетней. Максимальное значение относительного числа видов у более чем половины групп приходится на ст. 3 и в этом проявляется отличие от территориальной приуроченности таких же значений у абсолютного параметра. Пространственные изменения относительного числа видов происходят по тем же типам, что и абсолютного, но с сильным преобладанием «нормального» и особенно его «верхненормального» подтипа. Иными словами, относительное число видов в группах, по сравнению с абсолютным, проявляет себя как более устойчивый признак. На ИС с учетом величины относительного числа видов в МФП господствуют ведущие (64%) и однолетние (71%) водоросли. В этом наблюдается сходство с экологическим спектром сообщества на ЕС. Кроме того, среди Ch и Осh, исходя из величины относительного числа видов, наибольшее представительство получают те же группы, что и на ЕС. Неизменным остается и высокое сходство базовой части экоспектров Ch и Rh. Отличие касается пропорций мезо- и олигосапробионтов, морской и солоноватоводно-морской групп. Кроме этого, у Rh в МФП наблюдается синхронное доминирование морских и солоноватоводно-морских видов. Следует отметить, что значение индекса сапробности (0,07), рассчитанного по данным о сапробном составе фитоценозов, свидетельствует о существенном загрязнении вод прибрежья бухты Голландия. Сравнительный анализ таких показателей пространственной изменчивости числа видов в экогруппах на разных субстратах, как лимиты и тип вариации, а также среднее значение самих характеристик, показал следующее. Независимо от типа субстрата лимиты вариации абсолютного числа видов в одноименных группах чаще совпадают или близки к этому. Лишь у солоноватоводно-морской и однолетней групп крайние значения абсолютного числа видов на ИС гораздо выше, чем на ЕС. Сходство МФП и МФБ проявляется и на уровне среднего числа видов в морской, солоноватоводной, ведущей, редкой, олиго- и полисапробной группах. В других случаях существует различие разной степени. На ИС, по сравнению с ЕС, среднее абсолютное число видов выше в таких группах, как солоноватоводно-морская, сопутствующая, однолетняя и мезосапробная. На ЕС только у многолетников этот показатель больше. В обоих сообществах на разных твердых субстратах изменчивость абсолютного числа видов в группах проявляется однотипно, с преобладанием «нормы». При этом вклад изменчивости, превышающей «норму», на ЕС вдвое больше, чем на ИС. Возможно это связано с тем, что на ИС складываются топически более однородные условия обитания водорослей, чем на ЕС. Лимиты вариации относительного числа видов, подобно абсолютному, на разных субстратах в большинстве случаев совпадают. Это же касается и среднего относительного числа видов в солоноватоводной, редкой и олигосапробной группах. Такой же показатель в морской, ведущей и многолетней группах выше на ЕС, а в солоноватоводно-морской, сопутствующей, однолетней и мезосапробной – на ИС. Тип изменчивости относительного числа видов в половине групп на всех субстратах одинаков. У другой половины на ИС она не так интенсивна, как на ЕС. В целом, относительное число видов одноименных групп на разных субстратах меньше подвержено пространственным флуктуациям, чем абсолютное. Абсолютная и относительная фитомасса, доминанты МФБ и МФП бухты Абсолютная фитомасса ценоза МФБ на ЕС варьирует от 4,3 до 20,0 кг∙м-2 и в среднем для станции равна 11,9±3,8 кг∙м-2. Ее пространственная изменчивость по шкале Г.Н.Зайцева соответствует «верхней» норме. Максимум показателя приходится на ст. 4, минимум – на ст. 6. На ст. 7 абсолютная фитомасса ценоза близка к максимальной. Соотношение суммарной фитомассы видов трех отделов на ЕС свидетельствует о количественном преимуществе Осh. Вторая и третья позиции заняты Ch и Rh. Анализ распределения абсолютной фитомассы отделов на конкретных участках берега показывает случаи нарушения этого соотношения. Так, на ст. 3 и 7 фитомасса Ch превышает этот показатель для Осh более, чем в 1,5 раза. Кроме того, на последней станции у выхода из бухты фитомасса Осh и Rh совпадает. Пространственная изменчивость суммарной абсолютной фитомассы видов каждого отдела осуществляется по-разному. У Ch на участке от ст. 1 до ст. 3 наблюдается увеличение фитомассы, затем снижение и очередное повышение на ст. 7, но без достижения прежнего максимума. Все изменения данной характеристики Ch относятся к «большим» (Сv = 69%). У Осh они ближе к колебательным и «значительным». Rh – единственный отдел, абсолютная фитомасса которого возрастает от ст. 1 до ст. 7, а ее крайние значения различаются на порядки. Минимум этого показателя у МФБ и его отделов, как правило, приходятся на одну и ту же ст. 6, реже – на ст. 1. Максимумы зафиксированы на станциях, ограничивающих пляж, и единично – за причалом катеров. Наибольший размах значений фитомассы обнаружен у Осh, наименьший – у Rh. На ИС средняя абсолютная фитомасса МФП и составляющих его отделов макроводорослей ниже, чем на ЕС. Особенно это касается общей фитомассы видов Осh и всего ценоза, у которых разница составляет 2,7 и 1,6 раза. Средняя фитомасса Ch на ИС достигает 57% фитомассы ценоза, тогда как на ЕС такой же уровень характерен Осh. Минимум фитомассы Ch, Осh и всего МФП приходится на ст. 7 за причалом катеров, у Rh – на ст. 3. Максимум показателя у МФП, Ch, Осh зафиксирован на ст. 2, у Rh – на ст. 7, где таким же уровнем фитомассы отдел обладает в составе МФБ. На ИС самый большой размах значений абсолютной фитомассы у Ch (отличие от МФБ), самый низкий – у Rh (сходство с МФБ). Пространственные вариации показателя у Ch на обоих субстратах являются сильно выраженными, тогда как у Осh и всего МФП они не столь существенны, как в МФБ. Следует отметить и менее интенсивный характер пространственной изменчивости абсолютной фитомассы Rh на ИС. В целом, по средним показателям абсолютной фитомассы в МФБ лидируют Осh, в МФП их место занято видами Ch. На твердых субстратах бухты Голландия роль Rh как продуцентов незначительна. Крайние значения данного показателя на ЕС и ИС зафиксированы на неодноименных станциях. Средние значения фитомассы Осh и всего фитоценоза выше на ЕС, у других отделов эта разница не так заметна. Изменчивость абсолютной фитомассы Ch на субстратах одинаково «большая», у Осh и всего фитоценоза она значительнее на ИС, у Rh – на ЕС. Размах значений абсолютной фитомассы Ch на ИС превышает таковой на ЕС, то есть там, где уровень данного показателя у ценоза и его компонентов, наоборот, выше. На ЕС относительная фитомасса Ch варьирует от 19 до 71% и в среднем достигает 37±13%. Минимум и максимум показателя приурочены к ст. 1 и 3 соответственно. Пространственные изменения скорее происходят в колебательном режиме и по силе соответствуют «значительным». У Осh крайние значения относительной фитомассы выше, чем у Ch, однако размах их вариаций ниже (54%). Максимум и минимум показателя приурочены к тем же станциям, что и у Ch, но в обратном порядке: на станцию с высоким уровнем относительной фитомассы у зеленых водорослей приходится ее же низкий уровень у бурых и наоборот. Средняя относительная фитомасса Осh в 1,5 раза выше, чем у Ch, изменения по станциям происходят в том же режиме, в пределах «верхней» нормы. У Rh крайние значения, размах вариаций и среднее значение относительной фитомассы в несколько раз ниже, чем у других отделов. Иными выглядят сила и общая тенденция изменений относительной фитомассы Rh в пространстве. Их сила является «аномально» высокой (Сv = 134%). На ст. 1–4 относительная фитомасса одинаково мала (1% фитомассы ценоза), но, начиная со ст. 5, наблюдается ее многократное увеличение вплоть до своего максимума на ст. 7 (28%). В итоге, на ЕС средняя относительная фитомасса у Осh самая большая и более константная, самая низкая и лабильная – у Rh. У Ch и Осh изменения осуществляются в колебательном режиме. Станции с минимумом показателя частично совпадают, чего не скажешь о территориальной приуроченности максимумов. На ИС сообщество макроводорослей характеризуется существенным отличием не только на уровне относительной фитомассы, но и благодаря особенностям ее пространственной динамики. Так, размах вариаций доли Rh и Ch в общей фитомассе, а также ее среднее значение в 1,4-3 раза выше на бетонных плитах, чем на камнях. Первый параметр показателя у Осh на обоих субстратах одинаков. Второй параметр фитомассы этих водорослей в МФП вдвое ниже, чем в МФБ. Изменчивость доли фитомассы Ch на плитах ниже, Осh – выше, чем на камнях, а у Rh она одинаково «аномально» высокая. Лимиты вариации относительной фитомассы у Ch и, частично, у Rh на двух субстратах приходятся на одноименные станции. У Осh такая ситуация характерна только для максимума. Группа видов-доминантов МФБ с высокой абсолютной фитомассой включает C. barbata, C. sericea (Huds.) Kutz., U. intestinalis (L.) Nees. У первого вида самый большой индекс доминирования (86%). Затем следуют U. intestinalis (25%) и C. sericea (14%). На ИС среди доминантов вместо U. intestinalis находится Ceramium virgatum Roth. c индексом доминирования в 25%. Величина этого индекса у C. barbata здесь ниже, а у C. sericea достигает 50%. Содоминирующие виды на ЕС встречаются повсюду, кроме ст. 1. В ее состав наиболее часто входит Ulva rigida C. Ag., реже – C. barbata и Ceramium diaphanum (Lightf.) Roth. На ИС содоминанты присутствуют только на ст. 1 и представлены C. sericea. Применение шкалы Е.Л. Любарского позволило распределить виды на группы с разным уровнем относительной фитомассы. МФБ и МФП в этом отношении проявляют как сходство, так и различие. Их сходство невелико и касается господства малозначимых видов с низкой относительной фитомассой. Общим является и наличие среди субдоминантов U. intestinalis. В МФБ, наряду с малозначимыми видами, весомую роль в продукционном процессе выполняют и второстепенные виды. Вклад фитомассы субдоминантов в МФБ примерно вдвое ниже, чем в МФП, где их видовой состав частью не только другой, но и разнообразнее на один вид. Доминанты, не характерные для МФБ бухты, своим функционированием на ИС обеспечивают 14% общей фитомассы.
Заключение В ходе гидроботанических исследований донной растительности прибрежья бухты Голландия были идентифицированы 15 видов макроводорослей из отделов Ch, Осh и Rh. По таксономическому разнообразию лидирует Ch, что является отличительной чертой фитобиоты данного района. Флористическая структура фитоценозов на ИС и ЕС во многом идентична. Пространственные изменения видового состава МФБ по шкале вариабельности признаков относятся к «верхненормальным». Установлен факт увеличения видового разнообразия с удалением от зоны пляжа к выходу из бухты. Среднее число видов МФП, а также его Ch и Rh выше, чем в МФБ. Их пиковые значения на разных субстратах не всегда территориально совпадают. Водоросли бухты представлены видами 11 экологических групп, среди которых преобладают однолетние, ведущие, солоноватоводно-морские и мезосапробные виды. Доминирование двух последних групп является характерной чертой фитобиоты района исследования. Общая доля солоноватоводно-морских и солоноватоводных видов, величина индекса сапробности свидетельствуют о распреснении и загрязнении воды в бухте. Изменения абсолютного числа видов в группах на разных субстратах происходят однотипно. Относительное число видов в одноименных группах, но на разных субстратах подвержено пространственным вариациям меньше. Средняя абсолютная фитомасса МФП и входящих в него отделов ниже, чем у МФБ. По данному показателю в МФБ превалируют виды Осh, а в МФП – Ch. Доминанты с высокой абсолютной фитомассой в МФБ представлены C. barbata, C. sericea, U. intestinalis, в МФП место последнего вида занято C. virgatum. На любом твердом субстрате преобладают малозначимые виды с низкой относительной фитомассой.
Список литературы 1. Евстигнеева И.К., Танковская И.Н. Донная растительность антропогенно-нагруженных акваторий Черного моря // Вестник СевГТУ. 2004. Вып.55. С. 126–134. 2. Евстигнеева И.К. Фитомасса и доминантный комплекс видов в сообществах прибрежного экотона Крыма // Альгология. 2006. Т.16, №3. C. 325–336. 3. Зайцев Г.Н. Математика в экспериментальной ботанике. – М: Наука, 1990. – 296 с. 4. Зинова А.Д. Определитель зеленых, бурых и красных водорослей южных морей СССР. – М.-Л.: Наука, 1967. – 397 с. 5. Калугина А.А. Исследование донной растительности Черного моря с применением легководолазной техники // Морские подводные исследования. – М., 1969. – С.105–113. 6. Калугина-Гутник А.А. Изменения в донной растительности Севастопольской бухты за период с 1967 по 1977 гг. // Экология моря. 1982. Вып. 9. С. 48–61. 7. Калугина-Гутник А.А. Изменение видового состава фитобентоса в бухте Ласпи за период 1967-1983 гг. // Экология моря. 1989. Вып.31. С. 7–12. 8. Калугина-Гутник А.А. Фитобентос Черного моря. – Киев: Наук. думка, 1975.– 248 с. 9. Розенберг Г.С. Количественные методы экологии и гидробиологии // Cб. научн. трудов, посвященный памяти А.И. Баканова. – Тольятти: СамНЦ РАН, 2005. – 404 с. 10. Совга Е.Е., Мезенцева И.В., Хмара Т.В., Слепчук К.А. О перспективах и возможностях оценки самоочистительной способности акватории Севастопольской бухты // Экологическая безопасность прибрежной и шельфовой зон и комплексное использование ресурсов шельфа. 28. – Севастополь: ЭКОСИ-Гидрофизика, 2014. – С. 153–164. 11. Guiry M. D., Guiry G. M. AlgaeBase. World-wide electronic publication, National University of Ireland, Galway [Electronic resource]. – http: // www.algaebase.org. – (дата обращения – 07.05.2016). Статья поступила в редакцию 20.12.2016
Species’ composition, ecological structure and quantitative characteristics of the Gollandiya bay macroalgae Irina K. Evstigneeva, Irina N.Tankovskaya The species composition and ecological structure of macrophytobenthos and macrophytoperiphyton of the black-sea Gollandiya bay in the location of Training Centre for Marine Researches and Technology of Sevastopol State University were described for the first time. Ecology-taxonomic leaders and principal producers were identified. The main tendencies in spatial changes of macrophytobenthos and macrophytoperiphyton of the bay were analysed. Key words: Black Sea; Gollandiya bay; macrophytobenthos; macrophytoperiphyton; occurrence; ecology-taxonomic structure; phytomass; variability.
Об авторах Евстигнеева Ирина Константиновна - Evstigneeva Irina K. кандидат биологических наук Logrianin@nm.ru Танковская Ирина Николаевна - Tankovskaya Irina N. младший научный сотрудник, Институт морских биологических исследований имени А.О.Ковалевского РАН (ФГБУН ИМБИ), Севастополь, Россия (Federal State Institution of Science «The A.O.Kovalevsky Institute of Marine Biological Research of RAS» IMBR, Sevastopol, Russia) Logrianin@nm.ru Корреспондентский адрес: Россия, 299011, Севастополь, пр. Нахимова, 2, ФГБУН ИМБИ; тел. 88692550795.
ССЫЛКА НА СТАТЬЮ: Евстигнеева И.К., Танковская И.Н. Видовой состав, экологическая структура и количественная характеристика макроводорослей бухты Голландия (Черное море) // Вопросы современной альгологии. 2017. № 1 (13). URL: http://algology.ru/1127 Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно. При перепечатке ссылка на сайт обязательна
К разделу ОБЗОРЫ, СТАТЬИ И КРАТКИЕ СООБЩЕНИЯ 
|
|||
|
| ||