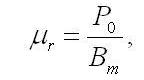|
|
|
|
по Материалам Международной конференции «Экологическая физиология водных фототрофов: распространение, запасы, химический состав и использование» V Сабининские чтения 29 ноября 2016 - 29 января 2017 Влияние освещенности на продуктивность и удельную скорость поддержания Arthrospira platensis Gomont
Авсиян А.Л.,Тренкеншу Р.П. Anna L. Avsiyan, Rudolf P. Trenkenshu
Институт морских биологических исследований им. А.О. Ковалевского (г. Севастополь)
УДК 582.232:581.121
Предложены простые уравнения, позволяющие количественно описать снижение продуктивности и плотности культуры микроводорослей после достижения стационарной фазы роста. Уравнения учитывают эндогенный расход биомассы при лимитировании роста светом. Приведены экспериментальные данные и результаты оценки влияния освещенности на продуктивность и удельную скорость поддержания Arthrospira platensis. Ключевые слова: микроводоросли; освещенность; продуктивность; удельная скорость поддержания; Arthrospira platensis.
Введение Современные исследования показывают, что как в темноте, так и во время освещения в клетках микроводорослей параллельно с фотосинтетическими реакциями также происходят и дыхательные процессы (Cournac et al., 2002, Raven, Beardall, 2016), приводящие к эндогенному расходу биомассы. Как и у высших растений, дыхание микроводорослей традиционно разделяют на «дыхание роста» и «дыхание поддержания». «Дыхание роста» обеспечивает необходимой энергией такие процессы, как поглощение неорганических ионов, включение этих ионов в промежуточные метаболические соединения и синтез из этих соединений структурных и функциональных компонентов клетки. Метаболические затраты на поддержание по определению не зависят от скорости роста, тогда как затраты на синтез отражают дополнительный прирост потребления энергии, необходимый для производства биомассы (Geider, Osborne, 1989; Amthor 2000). Потребность в энергии поддержания можно представить как расход биомассы в ходе эндогенных процессов. Удельная скорость поддержания может рассматриваться как скорость оборота биомассы. Это удобный показатель для сравнения скорости оборота компонентов биомассы и потребности в энергии, расходуемой на механизмы поддержания (Перт, 1978; Pirt, 1982). Соотношение интенсивности дыхательных процессов с интенсивностью фотосинтеза и скоростью роста давно вызывает большой интерес исследователей (Ryther, 1953; Geider, Osborne, 1989; Duarte, 1995), поскольку оно важно для моделирования роста и расчета продуктивности микроводорослей. Интенсивность дыхания микроводорослей зачастую оказывается линейно зависимой от их скорости роста (Geider, Osborne, 1989). В предлагаемой работе представлена попытка количественно оценить влияние освещенности на продуктивность и удельную скорость поддержания плотной культуры микроводорослей с помощью полуэмпирической модели.
Материалы и методы В экспериментальной работе в качестве объекта использовали прокариотическую микроводоросль Arthrospira platensis (штамм IMBR-31) из коллекции культур ИМБИ РАН. Культивирование осуществляли в накопительном режиме на питательной среде Заррук (Zarrouk, 1966) в культиваторах плоско-параллельного типа с рабочей толщиной 5 см. Объем питательной среды составлял 3 л. Этот объем поддерживали на протяжении всего эксперимента, доливая перед измерениями дистиллированную воду до метки. Температура культуральной среды составляла 35±2°С. Перемешивание осуществлялось путём непрерывного барботирования воздухом. Культиваторы непрерывно освещались лампой ДРЛ-700 с одной стороны и были закрыты светонепроницаемой пленкой с другой. В опыте было шесть вариантов с освещенностью рабочей поверхности культиватора от 2,5 до 16,2 кЛк. Освещенность на поверхности культиватора измеряли при помощи люксметра Ю-116. Отбор проб для определения плотности культуры проводили ежедневно. Прирост биомассы измеряли по изменению оптической плотности суспензии на фотоколориметре КФК-2 на длине волны 750 нм. Переход от единиц оптической плотности (D750) к величине абсолютно сухой массы (АСМ) осуществляли посредством эмпирического коэффициента k = 0,624 ± 0,049 г • л-1 • ед.опт.пл.-1, ACМ = k • D750.
Результаты и обсуждение Рассмотрим процессы, определяющие изменение продуктивности с увеличением плотности культуры низших фототрофов. Для каждой интенсивности света в данных условиях существует некоторое значение валовой продуктивности P0. Если вся падающая световая энергия поглощается клетками микроводорослей (как происходит в плотных культурах), то для неизменных условий и отсутствия лимитирования по другим факторам эта величина будет константой (P0 = const). Валовая продуктивность отражает скорость синтеза биомассы в клетке. Однако в то же время происходит эндогенный расход биомассы для энергетического обеспечения жизнедеятельности клетки. Скорость потери биомассы путем эндогенного расхода обозначим Pr:
где μr – удельная скорость поддержания, ч-1; B – плотность культуры, г • л-1. Наблюдаемая продуктивность P в каждый момент времени будет равна разности между валовой продуктивностью и скоростью эндогенного расхода биомассы:
где P0 – валовая продуктивность, г • л-1 • ч-1; Pr – скорость эндогенного расхода биомассы, г • л-1 • ч-1. Снижение величины продуктивности при росте плотности культуры обусловлено тем, что при неизменной валовой продуктивности возрастают потери, связанные с эндогенным расходом биомассы. После того, как накопительная культура микроводорослей достигает своей максимальной продуктивности, при повышении плотности культуры разность между максимальной продуктивностью и текущей будет равна разности в плотности, умноженной на удельную скорость поддержания:
где Pm – максимальная продуктивность г • л-1 • ч-1; B1 – плотность культуры при максимальной продуктивности, г • л-1. Отсюда можно выразить удельную скорость поддержания: одуктивности возрастают потери, связанные с эндогенным расходом биомассы. После того, как накопительная культура микроводорослей достигает своей максимальной продуктивности, при повышении плотности культуры разность между максимальной продуктивностью и текущей будет равна разности в плотности, умноженной на удельную скорость поддержания:
Стоит ещё раз отметить, что зависимости отражают соотношение плотности культуры и продуктивности только при таких плотностях, когда вся падающая энергия поглощается клетками, а также не оказывают лимитирующего влияния другие факторы. В накопительной культуре это происходит на стадии замедления роста. Когда накопительная культура микроводорослей достигает стационарной фазы роста и максимальной плотности Bm, наблюдаемая продуктивность равна нулю, поскольку валовая продуктивность становится равной эндогенному расходу биомассы.
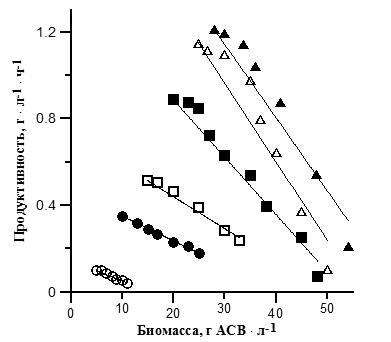
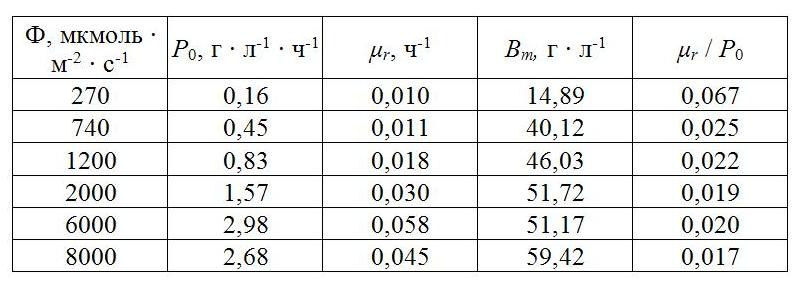
где Bm – максимальная плотность культуры, г • л-1. Исходя из литературных данных, можно отметить, что при плотности культуры выше оптимальной действительно наблюдается линейное снижение продуктивности (Hu et al., 1996; Hu et al., 1998). Используя эти данные (Hu, 1998), мы можем описать снижение продуктивности A. platensis с ростом биомассы при различной плотности потока фотонов (от 270 до 8000 мкмоль · м-2 · с-1), с помощью уравнения (1). На рис. 1 представлена зависимость продуктивности от плотности культуры при различной плотности потока фотонов (ППФ). Как видно из рис. 1, зависимость продуктивности от биомассы линейна и достоверно аппроксимируется полученным нами уравнением. Были определены коэффициенты этого уравнения (валовая продуктивность, удельная скорость поддержания). Расчетные данные представлены в табл. 1. Валовая продуктивность P0 линейно возрастала с ростом освещенности в диапазоне ППФ 270-2000 мкмоль • м-2 • с-1, однако при сверхвысоких ППФ 6000 и 8000 мкмоль • м-2 • c-1 продуктивность отклонялась от линейной зависимости из-за возможной фотодеструкции пигментов и ингибирования фотосинтеза в этих условиях. Удельная скорость поддержания тоже увеличивалась при увеличении освещенности, составляя от 0,01 до 0,58 ч-1. Примечательно, что в диапазоне ППФ 1200-6000 мкмоль • м-2 • с-1 отношение удельной скорости поддержания к валовой продуктивности изменялось незначительно (от 0,019 до 0,022). Очевидно, что величина удельной скорости поддержания зависит от валовой продуктивности и от скорости энергоемких синтетических процессов в клетке.
Рис. 1. Зависимость продуктивности A. platensis от плотности потока фотонов (ППФ; мкмоль•м-2•с-1) и плотности культуры. Фотобиореакторы были освещены с обеих сторон при следующей общей ППФ: ○ – 270; ● – 740; □ – 1200; ■ – 2000; ∆ – 6000; ▲ – 8000 мкмоль•м-2•с-1. Точки – данные (Hu, 1998), сплошные линии – аппроксимация данных уравнением (2). Fig. 1. A. platensis productivity as a function of photon flux density (PPD, μmol•m-2•s-1) and culture density. Photobioreactors were illuminated from both sides at total PPD: ○ – 270; ● – 740; □ – 1200; ■ – 2000; ∆ – 6000; ▲ – 8000 μmol•m-2•s-1. Symbols are data (Hu, 1998), solid lines are data approximation with equation (2).
Таблица 1. Ростовые характеристики A. platensis при различной ППФ Table 1. A. platensis growth characteristics at different PPD [based on data Hu et al. (1998)]
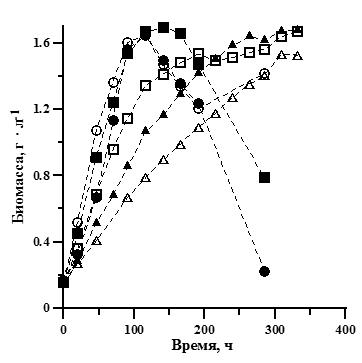
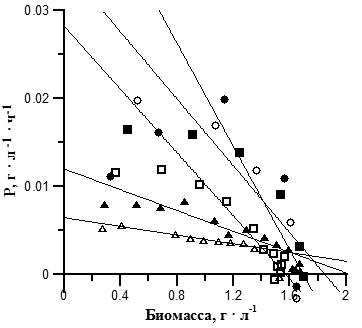
После анализа литературных данных нами был проведен эксперимент для выяснения закономерностей влияния освещенности на рост и эндогенный расход биомассы А. platensis (рис. 2).
Рис. 2. Накопительные кривые роста плотности культуры А. platensis при различной освещенности. Поверхностная освещенность: ∆ – 2,5; ▲ – 4; □ – 7,5; ■ – 11,5; ○ – 15; ● – 16,2 кЛк. Fig. 2. А. platensis batch cultures growth curves at different illumination. Illumination: ∆ – 2,5; ▲ – 4; □ – 7,5; ■ – 11,5; ○ – 15; ● – 16,2 klx.
Как видно из графиков на рис. 2, во всех вариантах опыта на накопительной кривой можно выделить фазу линейного роста, когда продуктивность постоянна и максимальна. Аппроксимацией данных на этом участке накопительной кривой мы можем определить максимальную продуктивность (Pm) (Тренкеншу, 2005):
где Bl – биомасса в начале линейной фазы, г • л-1; tl – время начала линейной фазы, ч. В вариантах с освещенностью 11,5–16,2 кЛк на 6–7 сутки эксперимента начиналась фаза отмирания. Наиболее резкое снижение плотности культуры наблюдалось в варианте с наибольшей освещенностью 16,2 кЛк. В вариантах с меньшими значениями освещенности (2,5-7,5 кЛк) снижение скорости роста происходило медленнее, с выраженной фазой замедления роста на накопительной кривой. Максимальная плотность культуры не различалась существенно при различных вариантах освещенности, но отличалось время достижения максимальной биомассы. На основании полученных данных был проведен расчет продуктивности для различных плотностей культуры, и на основании полученных графиков (рис. 3) выполнена аппроксимация по уравнению (1) для вычисления ростовых характеристик. Для варианта с максимальной освещенностью 16,2 кЛк вычисление не производилось из-за слишком короткой фазы замедления роста.
Рис. 3. Зависимость продуктивности А. platensis от плотности культуры при различной освещенности. Поверхностная освещенность: ∆ – 2,5; ▲ – 4; □ – 7,5; ■ – 11,5; ○ – 15; ● – 16,2 кЛк. Сплошные линии – аппроксимация данных уравнением (2). Fig. 3. A. platensis productivity as a function of culture density at different illumination. Illumination: ∆ – 2,5; ▲ – 4; □ – 7,5; ■ – 11,5; ○ – 15; ● – 16,2 klx. Solid lines are data approximation with equation (2).
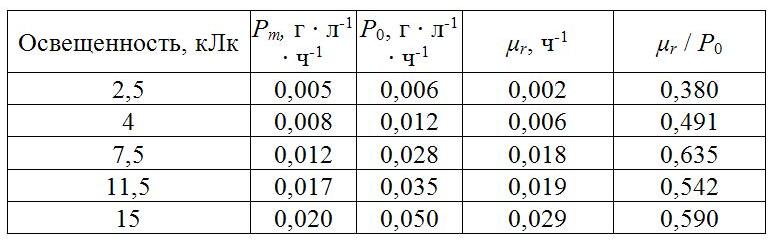
Рассчитанные ростовые характеристики А. platensis в накопительной культуре представлены в табл. 2. Максимальная наблюдаемая продуктивность линейно возрастала с увеличением освещенности от 0,005 до 0,02 г • л-1• ч-1, кроме варианта с наибольшей освещенностью 16,2 кЛк, при которой продуктивность составляла 0,017 г • л-1 • ч-1. Возможно, такое снижение продуктивности обусловлено фотоингибированием при одновременном действии высокой освещенности и высокой температуры (35±2°С).
Таблица 2. Ростовые характеристики А. platensis в накопительной культуре при различной освещенности
Остальные характеристики (валовая продуктивность, удельная скорость поддержания и их соотношение) рассчитывались только для вариантов с освещенностью 2,5–15 кЛк. Валовая продуктивность линейно возрастала с повышением освещенности, составляя от 0,006 до 0,05 г•л-1•ч-1, при этом зависимость хорошо описывается линейным уравнением вида P0 = k • I (R2=0,981). Удельная скорость поддержания также возрастала при увеличении поверхностной освещенности, изменяясь от 0,002 до 0,29 ч-1. Отношение удельной скорости поддержания к валовой продуктивности варьировало в небольших пределах, от 0,38 до 0,59. Значения удельной скорости поддержания в выполненном нами эксперименте имеют тот же порядок, что и значения, рассчитанные по данным (Hu, 1998). Так, μr = 0,018 для ППФ=1200 мкмоль • м-2 • с-1 по литературным данным, и μr = 0,019 для освещенности 11,5 кЛк по данным нашего эксперимента. Однако есть и некоторые различия при других значениях освещенности. В целом, выполненный анализ литературных и экспериментальных данных выявил сходные закономерности взаимосвязи плотности культуры с продуктивностью, освещенности с продуктивностью и удельной скоростью поддержания при культивировании цианопрокариоты А. platensis в накопительном режиме. Стоит отметить, что величина удельной скорости поддержания возрастает при увеличении освещенности, что скорее всего связано как с увеличением затрат энергии на обеспечение продукционных процессов, так и с увеличением интенсивности процессов фотодыхания, ведущих к рассеянию энергии.
Заключение Предложенный в работе подход позволяет вычислять валовую продуктивность и удельную скорость поддержания низших фототрофов, используя значения биомассы и продуктивности на фазе замедления роста в накопительной культуре. С помощью аппроксимации таких данных есть возможность вычислить удельную скорость поддержания и валовую продуктивность в культуре. Используя данный подход, проведена обработка литературных и экспериментальных данных для выявления влияния освещенности на ростовые характеристики А. platensis в накопительной культуре. Показано, что с увеличением освещенности происходит линейное возрастание наблюдаемой и валовой продуктивности, а также увеличение значения удельной скорости поддержания, которая может составлять до 0,029 ч-1.
Список литературы 1. Перт С.Дж. Основы культивирования микроорганизмов и клеток / Перевод с англ. под ред. Работновой И. Л. – М.: Мир, 1978. – 331 с. 2. Тренкеншу Р.П. Простейшие модели роста микроводорослей. 1. Периодическая культура // Экология моря. 2005. Т.67. С. 89–97. 3. Amthor J.S. The McCree – de Wit – Penning de Vries – Thornley respiration paradigms: 30 years later // Ann. Bot. 2000. 86. P. 1–20. 4. Cournac L., Latouch G., Cerovic Z. et al. In vivo interactions between photosynthesis, mitorespiration, and chlororespiration in Chlamydomonas reinhardii // Plant Phys. 2002. V.129, №4. P. 1921–1928. 5. Duarte P. A mechanistic model of the effects of light and temperature on algal primary productivity // Ecol. Model. 1995. V.82, Iss.2. P. 151–160. 6. Geider R.J., Osborne B.A. Respiration and microalgal growth: a review of quantitative relationship between dark respiration and growth // New Phytol. 1989. V.112. P. 327–341. 7. Hu Q., Guterman H., Richmond A. Physiological characteristics of Spirulina platensis (Cyanobacteria) cultured at ultrahigh cell densities // J. Phycol. 1996. V.32. P. 1066–1073. 8. Hu Q., Zarmi Y., Richmond A. Combined effects of light intensity, light-path and culture density on output rate of Spirulina platensis (Cyanobacteria) // Eur. J. Phycol. 1998. V.33. P. 165–171. 9. Pirt S.G. Maintenance energy: a general model for energy-limited and energy-sufficient growth // Arch. Microbiol. 1982. V.133, Iss.4. P. 300–302. 10. Raven J.A., Beardall J. Dark respiration and organic carbon loss // The Physiology of Microalgae. – Springer International Publishing, 2016. – P. 129–140. 11. Ryther J.H. The ratio of photosynthesis to respiration in marine plankton algae and its effect upon the measurement of productivity // Deep Sea Res. 1953. V.2, Iss.2. P. 134–139. 12. Zarrouk C. Contribution à l’étude d’une cyanophycée. Influence de divers facteurs physiques et chimiques sur la croissance et la photosyntèse de Spirulina maxima (Stech. Et Gardner) Geitler: Ph.D. thesis. – Paris, 1966. – 138 p. Статья поступила в редакцию 13.12.2016
Influence of illumination on productivity and specific maintenance rate Anna L. Avsiyan, Rudolf P. Trenkenshu Simple equations that allow describing quantitatively productivity and density decrease of the microalgae culture after reaching the stationary phase of growth are offered. The equations take into account endogenous biomass consumption at growth limiting by light. Experimental data and results of the assessment of illumination influence on productivity and specific maintenance rate of Arthrospira platensis are given. Key words: microalgae; illumination; productivity; specific maintenance rate; Arthrospira platensis.
Об авторах Авсиян Анна Львовна - Avsiyan Anna L. младший научный сотрудник, Институт морских биологических исследований им.А.О.Ковалевского РАН (ФГБУН ИМБИ), г. Севастополь, Россия (Federal State Institution of Science «The A. O. Kovalevsky Institute of Marine Biological Research of RAS» IMBR, Sevastopol, Russia), Отдел биотехнологий и фиторесурсов anna.l.avsiyan@gmail.com Тренкеншу Рудольф Павлович – Trenkenshu Rudolf P. кандидат биологических наук r.trenkenshu@rambler.ru Корреспондентский адрес: Россия, 299011, Севастополь, пр. Нахимова, 2, ФГБУН ИМБИ; тел. (869)-2550795.
ССЫЛКА НА СТАТЬЮ: Авсиян А.Л., Тренкеншу Р.П. Влияние освещённости на продуктивность и удельную скорость поддержания Arthrospira platensis Gomont // Вопросы современной альгологии. 2017. № 1 (13). URL: http://algology.ru/1107 Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно.
При перепечатке ссылка на сайт обязательна
К разделу ОБЗОРЫ, СТАТЬИ И КРАТКИЕ СООБЩЕНИЯ 
|
|||||||||||||
|
| ||||||||||||