|
|
|
|
V Сабининские чтения 29 ноября 2016 - 29 января 2017 Структурные перестройки микросообществ как мера устойчивости природных водных экосистем
Анциферова Г.А.1, Русова Н.И.2 Galina A. Antsiferova, Nadezhda I. Rusova
1Воронежский государственный университет
УДК 551.435.136
В бассейне среднего течения реки Ворона располагаются озера Кипец и Рамза. В течение ряда лет исследовалась структура микросообществ, представленных диатомовыми водорослями и цианобактериями. Как мера устойчивости водных экосистем рассматриваются структурные перестройки таксономического состава фитопланктона и микрофитобентоса. В качестве контроля функционирования экосистем на уровне сохранения экологического благополучия использован метод биоиндикации. Ключевые слова: биоиндикация; водная экосистема; диатомовые водоросли; сообщество микроводорослей; устойчивость водной экосистемы; функционирование водной экосистемы; цианобактерии; экология.
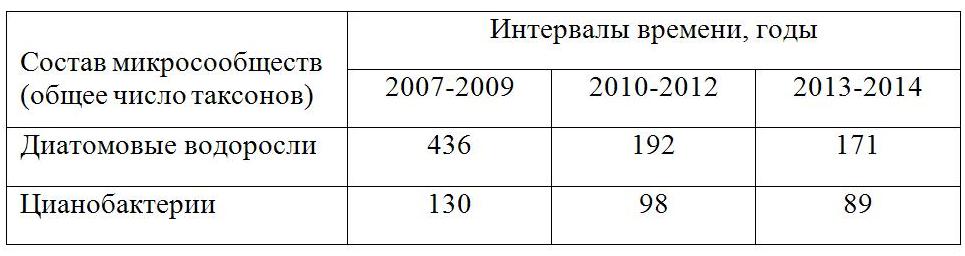
В бассейне среднего течения реки Ворона с 1994 года существует особо охраняемая природная территория, – заповедник «Воронинский». Озера Рамза и Кипец, которые представляют собой озеровидные расширения русла реки, входят в состав заповедника. Эти водные экосистемы являются мелководными и среднеглубокими хорошо прогреваемыми водоемами с широко развитыми зонами высшей водной растительности. По трофическому статусу, то есть по степени обеспеченности биогенными веществами, по соотношению процессов продуктивности, аккумуляции и деструкции органического вещества, озера относятся к эвтрофному типу. Поставщиками биогенов являются толщи слагающих водосборы мореных отложений донского оледенения, перекрывающие их проблематичные покровные суглинки и почвы зоны аэрации. Значительные объемы биогенных веществ связаны также с высокой степенью сельскохозяйственного освоения территории, ее распашкой и внесением удобрений. Исследования температурного режима вод р. Ворона показывают, что максимума 25,5–26,5ºС температуры достигают в июле месяце, минимальные температуры 0,2ºС наблюдаются с конца декабря до начала весеннего таяния снега. Средняя температура составляет 11,9ºС, устойчивый переход через +10ºС происходит весной в третьей декаде апреля, осенью – во второй декаде октября. Температура вод, их прозрачность, газовый и гидрохимический состав регулируют питание, фотосинтез, рост (деление) диатомовых водорослей. Температурный режим в водоемах средних широт в течение вегетационного сезона определяет два максимума развития диатомей: весна – первая половина лета и вторая половина августа – осенний период. Для теплолюбивых видов диатомей оптимальными являются температуры от 20 до 28ºС, для холодноводных видов – от 4–10 до 15ºС. Диатомовые водоросли начинают активно развиваться с начала апреля, и их вегетация прекращается в октябре – начале ноября, по мере охлаждения вод. В середине лета, с установлением постоянной температуры вод 23–25ºС и более, наблюдается максимум в развитии цианобактерий. В экологически благополучных водных экосистемах озер Рамза и Кипец в это время существуют условия, способствующие распространению вегетационной сукцессии цианобактерий, которую отличает разнообразный видовой состав. Отдельные виды достигают массового развития, однако «цветения» вод в том виде, как оно проявляется в загрязненных и грязных водоемах, не наблюдается. В водоемах заповедника реализуется свойство многих представителей цианобактерий, наряду с высшими водными растениями, диатомовыми водорослями и бактериями, включаться в общую систему процессов фотосинтеза, круговорота биогенных веществ, переработки органического вещества вплоть до его минерализации (Садчиков, Кудряшов, 2004). Проточно-русловой гидродинамический режим, морфометрические и морфологические параметры озерных акваторий создают широкий диапазон гидрофизических и гидрохимических условий. На фоне этого трудно переоценить значение обширных зон мелководий, заросших разнообразной водной растительностью. Речные озера Вороны характеризуются многообразием биотопов, благодаря чему наблюдается чрезвычайное видовое богатство сообществ диатомовых водорослей и цианобактерий (Анциферова, 2012). Изучение фитопланктона и микрофитобентоса водоемов заповедника в течение 2007–2014 гг. позволило выделить в развитии водорослей отдельные временные интервалы. 2007–2009 годы развития водных экосистем рассматриваются как интервал времени, отвечающий среднегодовым параметрам температур и осадков для территории лесостепной провинции Приволжской возвышенности (Русова, 2015а). Сообщества диатомовых водорослей и цианобактерий данного периода исследований отличает наибольшее число таксонов (табл. 1).
Таблица 1. Изменение общего числа таксонов в сообществах фитопланктона и микрофитобентоса по годам (2007–2014 гг.)
В составе сообществ планктонных диатомовых водорослей в целом широко распространены с оценками обилия «в массе», «очень часто», «часто» такие виды как Stephanodiscus hantzschii Grun., S. rotula (Kütz.) Hendey, Aulacoseira alpigena (Grun.) Krammer, А. italica (Ehr.) Grun. et var. tenuissima (Grun.) Sim., A. granulata (Ehr.) Sim., A. islandica ssp. helvetica (O.Müll.) Sim., Cyclostephanos dubius (Fricke) Round, Melosira varians Ag., Fragilaria crotonensis Kitt., Asterionella gracillis (Hantzsch.) Heib., Synedra berolinensis Lemm. В сообществах микрофитобентоса преобладают виды Fragilaria construens (Ehr.) Grun. et var. venter (Ehr.) Grun., F. brevistriata Grun., Tabellaria flocculosa (Roth.) Kütz., Eunotia lunaris (Ehr.) Grun., E. bigibba Ehr., Synedra ulna (Nitzsch.) Grun. et var. aequalis (Kütz.) Hust., S. tabulata (Ag.) Kütz., Navicula anglica Ralfs., N. bacillum Ehr., N. bicapitata Hust., N. cari Ehr., N. cuspidata Kütz. et var. ambigua (Ehr.) Grun. et var. hankae Skv. et f. craticularis Skv., N. cryptocephala Kütz., N. crucicula (W. Sm.) Donk., N. hungarica Grun. et var. capitata Cl., N. gastrum Ehr. et var. limnetica Skv., N. lacustris var. elongata Skv. et Mayer, N. oblonga Kütz., N. pupula Kütz. et var. rectangularis (Greg.) Grun, N. platystoma Ehr., N. placentula (Ehr.) Grun. et f. latiuscula (Grun.) Meist. et var. rostrata A. Mayer, N. pusio Cl., N. radiosa Kütz., N. tuscula (Ehr.) Grun., Gomphonema augur Ehr., G. acuminatum var. coronatum (Ehr.) W. Sm. et var. trigonocephala (Grun.) Cl., G. constrictum Ehr., G. intricatum Kütz. et var. pumilum Grun., G. olivaceum (Lyngb.) Kütz., G. sphaerophora Ehr., G. parvulum (Kütz.) Grun., Epithemia sorex Kütz. et var gracilis Hust., E. intermedia Fricke, E. zebra (Ehr.) Kütz. et var. porcellus (Kütz.) Grun. et var. saxonica (Kütz.) Grun., E. turgida (Ehr.) Kütz. et var. granulata (Ehr.) Grun., Cymbella ventricosa Kütz., C. leptoceros (Ehr.) Grun., Cymbella cistula (Hemp.) Grun. et var. maculata (Kütz.) V. H., C. turgida (Greg.) Cl., C. ventricosa Kütz., C. cymbiformis (Ag.? Kütz.) V. H., C. microcephala Grun., C. tumida (Bréb.) V.H., C. norvegica Grun., C. obtusiuscula (Kütz.) Grun., C. hybrida Grun., Stauroneis anceps Ehr. et f. gracilis (Ehr.) Cl., Achnanthes munitissima Kütz., Stauroneis anceps Ehr., Nitzschia acicularis W. Sm., Nitzschia subtilis (Kütz.) Grun., N sublinearis Hust., Cocconeis placentula Ehr. et var. euglypta (Ehr.) Cl. et var. intermedia (Herib. et Perag.) Cl. et var. rouxii (Brun. et Perag.) Cl., C. pediculus Ehr. et var. minutissima Poretzky, Denticula tenius Kütz., D. elegans Kütz., Rhopalodia gibba (Ehr.) O.Müll. et var. ventricosa (Ehr.) Grun., Amphora ovalis Kütz. et var. libyca Kütz. et var. gracilis Ehr., Gyrosigma acuminatum (Kütz.) Rabenh., G. attenuatum (Kütz.) Rabenh., G. scalproides (Rabenh.) Cl., Cymatopleura solea (Bréb.) W. Sm. et var. apiculata (W. Sm.) Ralfs. и многие другие. Среди цианобактерий повсеместно наблюдаются с оценками обилия «в массе», «очень часто», «часто», «нередко» Dactylococcopsis raphidioides Hansg., Merismopedia glauca f. insignis (Schkorb.) Geitl., Pseudoholopedia convoluta (Bréb.) Elenk., Microcystis hansgirgiana (Hang.) Elenk., M. pulverea (Wood) Forti emend. Elenk., M. pulverea f. holsatica (Lemm.) Elenk., M. wesenbergii Komarek, Gloeocapsa limnetica (Lemm.) Hollerb., Coelospherium kützingianum Näg., Anabaena hassalii f. minor V. Poljansk., A. spiroides Kleb., Nodularia spumigena Mert., Pseudoanabaena bipes Böcher, Ostillatoria planctonica Wolosz., Borzia sudetica Erceg., Spirulina abbreviata Lemm., S. massartii (Kuff.) Geitl., Romeria leopoliensis (Racib.) Koczw., Phormidium ambiguum Gom., Ph. foveolarum (Mont.) Gom., Ph. papyraceum (Ag.) Gom., Ph. uncinatum (Ag.) Gom., Lyngbya aestuarii (Mert.) Liebm., L. lagerheimii (Möb.) Gom., L. spendens Tschern., Schizothrix fragilis (Kütz.) Gоm. и многие другие. В период 2010–2012 годов возникли стрессовые для сообществ фитопланктона и микрофитобентоса условия среды обитания, обусловленные аномально высокими летними температурами воздуха, вызвавшими повышенный прогрев водной массы. Экстремально высокие летние температуры воздуха обусловили изменение температурных показателей региона (Акимов, Акимов, 2014; Акимов, Задорожная, 2014). Но при этом они позволили выявить чрезвычайную значимость данного природного фактора для функционирования водных экосистем. Высокие летние температуры воздуха способствовали тому, что температура речных вод поднималась до 22–23ºС и 27–28ºС. В озерных водоемах воды приповерхностных слоев в дневное время в июле – начале августа 2010–2012 гг. прогревались до температуры воздуха, часто достигая 37–40ºС. В результате кислородный режим в конце летнего периода изменялся до содержаний ниже ПДК, то есть менее 4 мг/л. Однако заморных явлений зарегистрировано не было (Летопись Природы …, 2013). В этот период произошло общее снижение видового богатства микросообществ (см. табл. 1). В сообществах планктонных диатомовых водорослей в целом широко распространены с оценками обилия «в массе», «очень часто», «часто» такие виды как Stephanodiscus hantzschii Grun., Cyclostephanos dubius (Fricke) Round., Cyclotella comensis Grun., C. kützingiana Thw, Thalassiosira lacustris (Grun.) Patrik., Aulacoseira italica (Ehr.) Sim, Melosira varians Ag., Synedra berolinensis Lemm. В сообществах микрофитобентоса преобладают виды Fragilaria brevistriata Grun., F. capucina Desm., F. construens (Ehr.) Grun. et var. venter (Ehr.) Grun., Synedra capitata Ehr., S. ulna (Nitzsch.) Grun. et var. aequalis (Kütz.) Hust. et var biceps (Kütz.) Schönf. et var. danica (Kütz.) Grun., et var impressa Hust. et var. spatulifera Grun., S. tabulata (Ag.) Kütz., Cocconeis pediculus Ehr., C. placentula Ehr., C. thumensis A. Mayer., Navicula gracilis Ehr., N. lanceolata (Ag.) Kütz., N. hungarica var. capitata Cl., N. oblonga Kütz., N. radiosa Kütz., Cymbella affinis Kütz., C. cistula (Hemp.) Grun. et var. maculata (Kütz.) V. H., C. cymbiformis (Ag.? Kütz.) V. H., C. hebridica (Greg.) Grun. C. hustedtii Krasske., C. lanceolata (Ehr.) V. H., C. ventricosa Kütz., Gomphonema acuminatum Ehr. et var. coronatum (Ehr.) W. Sm. et var brebissonii (Kütz.) Cl. et var. trigonocephala (Grun.) Cl., G. augur Ehr., G. constrictum Ehr. et var. capitatum (Ehr.) Cl., G. olivaceum (Lyngb.) Kütz. et var. calcarea Cl., G. parvulum (Kütz.) Grun., G. intricatum Kütz., Anomoeoneis sphaerophora (Kütz) Pfitz., Gyrosigma acuminatum (Kütz.) Rabenh., Amphora ovalis Ehr. et var. gracilis Ehr. et var. libyca Ehr. et var. pediculus Kütz., A. perpusilla Grun., Epithemia intermedia Fricke., E. sorex Kütz., E. mülleri Fricke., E. turgida (Ehr.) Kütz. et var. granulata (Ehr.) Grun., E. zebra (Ehr.) Kütz. et var. saxonica (Kütz.) Grun. et var. porcellus (Kütz.) Grun., Rhopalodia gibba (Ehr.) O. Müll. et var. ventricosa (Ehr.) Grun., Nitzschia kützingiana Hilse., N. gracilis Hantzsch. et var. capitata Wisl. et Por., N. sigmoidea (Ehr.) W. Sm. и другие. В сообществах цианобактерий повсеместно с оценками обилия «в массе», «очень часто», «часто», «нередко» наблюдаются Synechocystis pevalekii Elenk., Dactylococcopsis planctonica Teiling., Holopedia irregularis Lagerh., Merismopedia punctata Meyen., M. tenuissima Lemm., Pseudoholopedia convolutа (Bréb.) Elenk., Tetrapedia gothica Reinsch., Microcystis aeruginosa f. pseudofilamentosa (Crow) Elenk., Lemmermennia pallidа (Lemm.) Elenk., Coelosphaerium kützingiana Näg., Pleurocapsa minor Hansg. emend. Geitl., Chlorogloea purpurea Geitl., Calotrix kossinskajae V. Poljansk., Mastigocladus laminosus Gohn., Sphaeronostoc pruniforme (Ag.) Elenk., Nematonostoc fragelliformis (Berk. et Curt.) Elenk., Borzia trilocularis Cohn., Rivularia planctonica Elenk., Anabaena contorta Bachm., A. constricta (Szaf.) Geitl., A. minima Tschernov., A. viguieri Denis et Frémy., Aphanothece clatrata W. et G. S. West., Ostillatoria irrigua (Kütz.) Gom., O. lacustris (Kleb.) Geitl., O. ornata f. planctonica Elenk., O. planctonica Wolosz., O. tenius Ag., Spirulina minima Wurtz., S. okensis Meyer., S. tenuiоr (Lagerh.) Kirchn., S. laxissima G. S. West., S. major Kütz., S. meneghiniana Zanard., Phormidium crouanii Gom., Ph. fragile (Menegh.) Gom., Ph. tenue (Menegh.) Gom., Ph. ambiguum Gom., Ph. foveolarum (Mont.) Gom., Ph. laminosum (Ag.) Gom., Ph. molle (Kütz.) Gom., Ph. valderiae (Delp.) Geitl., Lyngbya aestuarii (Mert.) Liebm., L. birgei G.M. Shith., L. lacustris Lemm., L. lagerheimii (Möb.) Gom., L. kossinskajae Elenk., L. truncicola Ghose., L. spirulinoides Gom., Stygonema hormoides (Kütz.) Born. et Flah. и другие. В 2010-2012 гг. в составе микросообществ в водоемах заповедника встречены таксоны, ранее в регионе не известные. Например, среди диатомовых водорослей это Neidium dubium var. constricta Hust., Caloneis ignorata Skv., Amphora costulata Skv., Gomphonema ventricosum f. curta Skv., Diatoma elongatum var. capitellata Poretzky, Eunotia fallax var. gracillima Krasske, Cymbella ventricosa var. ovata Grun., Surirella didyma Kütz. Pinnularia rangoonensis Grun., Stauroneis wislouchii f. parva Poretzky et Anissimova. В составе сообществ цианобактерий это Merismopedia elegans A. Br., Gomphosphaeria lacustris f. compacta (Lemm.) Elenk., Woronichinia naegiliana (Ung.) Elenk., Spirulina labyrinthiformis (Menegh.) Gom., S. minima A. Wurtz., Anabaena minima Tschernov, Ostillatoria ornatа f. planctonica Ehr., Phormidium fragile (Menegh.) Gom., Stigonema hormoides (Kütz.) Born. et Flah., Beckia bella (G. Beck) Elenk., Microcoleus lacustris (Rabenh.) Farl., наблюдаются также ранее неизвестные в водоемах территории России и центральных районов европейской части виды, такие как Romeria chlorina Bőcher, Lyngbya majusculа Harvey, Spirulina curta (Lemm.) Geitl. и другие таксоны (Анциферова, 2013). Большой интерес вызывает распространение в заливах оз. Кипец с оценками обилия «в массе» вида Ostillatoria coerulescens Gicklh. Для массового распространения его зеленых дерновинок, имеющих «голубой блеск», благоприятны условия существования в иле, при выделении сероводорода. В 2012 г. выразительно проявились экологическая значимость и высокие миграционные свойства цианобактерий. В связи с повышением температурного режима произошло вселение и массовое развитие видов цианобактерий, чуждых для региона, характерных для хорошо прогреваемых водоемов, например, Бухары, Крыма, и вод горячих термальных источников (Анциферова, 2013; Русова, 2013). Резко увеличились сроки их активной вегетации (конец июня – август, и даже первая декада сентября), и общие объемы органического вещества, опускающегося на дно в конце сезона вегетации. Цианобактерии распространились повсеместно по акваториям озер, и не только в виде отдельных плавающих дерновинок и пленок на поверхности воды, но и плотно облекая стебли высших водных растений, например, рдестов, роголистника, урути и других макрофитов. Массовое развитие цианобактерий позволило проследить их адаптационные возможности, проявившиеся в эти, а также в последующие 2013-2014 годы. В период 2013–2014 годов температурные показатели вернулись к среднестатистическим значениям. Однако таксономическая структура микросообществ не восстановилась в рамках 2007–2009 годов. Общее видовое богатство микросообществ еще более снизилось (см. табл.). В сообществах планктонных диатомовых водорослей в целом широко распространены с оценками обилия «в массе», «очень часто», «часто» такие виды как Thalassiosira lacustris (Grun.) Hasle., Cyclotella meneghiniana Kütz., Aulacoseira granulata (Ehr.) Sim., Aulacoseira distans (Ehr.) Sim., A. italica (Ehr.) Sim., Melosira varians Ag., Synedra berolinensis Lemm. В сообществах микрофитобентоса преобладают виды Ellerbeckia arenaria (Moore ex Ralfs) Grawford., Fragilaria brevistriata Grun., F. capucina Desm., F. construens (Ehr.) Grun. et var. venter (Ehr.) Grun F. crotonensis Kitt., F. pinnata Ehr., Synedra amphicephala Kütz., S. capitata Ehr., S. ulna (Nitzsch.) Grun. et var. biceps (Kütz.) Schönf. et var. spatulifera (Kütz.) Hust., Rhoicosphenia curvata (Kütz.) Grun., Eucocconeis elliptica Saveljewa-Dolgowa., Diatoma anceps (Ehr.) Kirchn., Eunotia faba (Ehr.) Grun., E. monodon (W. Sm.) Hust., E. praerupta Ehr., Cocconeis pediculus Ehr., C. placentula Ehr., C. thumensis A. Mayer., Navicula cryptocephala et Kütz., N. cuspidata Kütz., N. gracilis Ehr., N. lanceolata (Ag.) Kütz. et var. tenuirostris et Skv., N. oblonga Kütz., N. hasta Pant., N. pupula Kütz., N. radiosa Kütz., N. menisculus Schum., N. hungarica var. capitata Cl., N. verecunda Hust., Caloneis silicula (Ehr.) Cl., Anomoeoneis sphaerophora (Kütz) Pfitz., Pinnularia viridis (Nitzsch.) Ehr., Cymbella affinis Kütz., C. aspera (Ehr.) Cl., C. cymbiformis (Ag.? Kütz.) V. H., C. cistula (Hemp.) Grun. et var. maculata (Kütz.) V. H., C. lanceolata (Ehr.) V. H., Cymbella leptoceros (Ehr.) Grun., C. hustedtii Krasske., C. hebridica (Greg.) Grun., C. prostrata (Berk.) Cl., C. tumida (Bréb.) V. H., C. turgidula (Greg.) Cl., C. ventricosa Kütz., Epithemia sorex Kütz., Epithemia intermedia Fricke., E. turgida (Ehr.) Kütz. et var. granulata (Ehr.) Grun., E. zebra (Ehr.) Kütz. et var. porcellus (Kütz.) Grun. et var. saxonica (Kütz.) Grun., Rhopalodia gibberula (Ehr.) O. Müll., Rhopalodia gibba (Ehr.) O. Müll. et var. ventricosa (Ehr.) Grun., Rh. parallela Grun., Gomphonema acuminatum Ehr. et var. coronatum (Ehr.) W. Sm. et var. brebissonii (Kütz.) Cl. et var. trigonocephala (Grun.) Cl., Gomphonema augur Ehr., Gomphonema constrictum Ehr. et var. capitatum (Ehr.) Cl., Gomphonema longiceps Ehr. et var. subclavatum Hust., G. olivaceum (Lyngb.) Kütz., G. parvulum (Kütz.) Grun., Achnanthes lanceolata (Bréb.) Grun. et var. capitata O. Müll., Denticula tenius Kütz., Gyrosigma acuminatum (Kütz.) Rabenh., G. attenuatum (Kütz.) Rabenh., Amphora ovalis Ehr. et var. gracilis et var. libyca Ehr. et var. pediculus Kütz., A. perpusilla Grun., A. veneta Kütz., Cymatopleura solea (Bréb.) W. Sm., Nitzschia holsatica Hust., N. linearis W. Sm., N. dissipata Kütz., N. kützingiana Hilse., N. sigma (Kütz.) W. Sm., N. sigmoidea (Ehr.) W. Sm. и другие. В сообществах цианобактерий повсеместно с оценками обилия «в массе», «очень часто», «часто», «нередко» наблюдаются Synechocystis aequatilis Sauv., Xenococcus chroococcoides F. E. Frisch., Merismopedia glauca (Ehr.) Näg., M. tenuissima Lemm., Pleurocapsa minor Hansg. emend. Geitl., Microcystis pulverea (Wood) Forti emend. Elenk., Chlorogloea purpurea Geitl., Tetrarcus ilsteri Skuja., Rabdoderma lineare Schmidle et Laut. emend. Hollerb., Gomphosphaeria lacustris f. compacta (Lemm.) Elenk., Anabaena constricta (Szaf.) Geitl., A. contorta Bachm., A. solitaria Klebs., Aphanothece castagne et (Bréb.) Rhabenh., A. clatrata W. et G. S. West., A. clatrata var. brevis Bachm., Coelosphaerium kützingiana Näg., Nodularia spumigena et Mert., N. spumigena f. luterea (Kütz.) Elenk., Rivularia rufescens (Näg.) Born. et Flah., Ostillatoria coerulensis Gicklh., O. irrigua et (Kütz.) Gom., O. lauterbornii Schmidle., O. ornara f. planctonica Elenk., O. lacustris et (Kleb.) Geitl., O. planctonica Wolosz., O. putrida Schmidle., O. utermoehliana (Utern.) Elenk., Spirulina meneghiniana Zanard., S. curta (Lemm.) Geitl., S. laxissima G. S. West., S. tenuissima Kütz., S. jenneri (Hass.) Kütz., Phormidium ambiguum Gom., Ph. papyraceum (Ag.) Gom., Ph. foveolarum (Mont.) Gom., Ph. frigidum F. E. Fritsch., Ph. mucicola Hub.-Pestalozzi et Naum., Ph. tenue (Menegh.) Gom., Lyngbya aestuarii (Mert.) Liebm., L. birgei G.M. Shith., L. lagerheimii et (Möb.) Gom., L. majuscula Harvey., L. truncicola Ghose. Из состава сообществ цианобактерий в 2013–2014 гг. исчезли виды горячих и теплых вод. Среди представителей теплолюбивых видов остаются те, которые наблюдались и ранее, в 2007-2009 гг. Но важной особенностью состава сообществ цианобактерий явилось сохранение и более широкое распространение таксонов-индикаторов загрязненных местообитаний, что отразило общее увеличение объемов отмирающего органического вещества. Среди них получили распространение такие виды как Rhabdoderma lineare f. compositum (G. V. Smith) Hollerb., Microcystis wesenbergii Komarek, Anabaena affinis Lemm., Aphanothece clatrata W. et G. S. West, A. stagnina (Spreng.) B.-Peters. et Geitl., Stigonema informe Elenk., Calothrix parietina (Näg.) Thur., Cloeotrichia echinulata (J. S. Smith.), Ostillatoria splendida Grev., O. tenius Ag., Schizothrix calcicola (Ag.) Gom., Seguenzaea sicula Borzi, Amorphonostoc paludosum (Kütz.) Elenk., Wollea saccata (Wolle) Born. et Flah. и другие. В 2013–2014 гг. установлено, что на отдельных участках акваторий в придонных слоях получили распространение полисапробные воды (Русова, 2013, 2015б). Это явилось следствием того, что стрессовые температурные условия способствовали образованию бόльших объемов органического вещества, при недостаточности возможностей его полной переработки и минерализации. Органическое вещество образуется как за счет отмершей высшей водной растительности, так и массовых скоплений опускающихся на дно в конце вегетации остатков цианобактерий, которые не успевают полностью разрушиться, формируя вторичное загрязнение вод. В целом структурные перестройки сообществ микроводорослей, как отклик на изменившиеся природные условия, направлены на поддержание устойчивости водных экосистем, поскольку микроводоросли непосредственно участвуют в процессах самоочищения вод и формирования их качества. Исследования таксономического состава сообществ диатомовых водорослей и цианобактерий по временным интервалам позволило проследить динамику их структурных перестроек, спровоцированные резкими температурными аномалиями 2010–2012 гг., при этом в 2013–2014 гг. структура сообществ также изменилась по сравнению с 2010–2012 гг., но не восстановилась в рамках 2007–2009 гг. Интенсивное обмеление и зарастание среднеглубоких и мелководных водоемов вызвало изменения в структуре сообществ высших водных растений, не только появились новые экологические ниши, но часть их перестала существовать. Это также повлияло на условия местообитания микросообществ. На экстремально высокие летние температуры воздуха и вод диатомовые, в связи с их температурной приуроченностью к умеренно-теплым условиям, отреагировали уменьшением таксономического состава сообществ (в сравнении с 2007–2009 гг.). Отдельные же представители цианобактерий в этих условиях проявили высокие миграционные свойства. Именно они в течение трех лет периода 2010–2012 гг. составили новую, оригинальную структуру вегетационной сукцессии. Произошел процесс вселения и массового распространения видов цианобактерий, чуждых для региона, в том числе видов-индикаторов загрязнения. В дальнейшем, в 2013–2014 гг., при переходе температурного режима к среднестатистическим для территории исследований показателям, многие из них выпали из состава сообществ. Реакция сообществ фитопланктона и микрофитобентоса на изменившиеся природные условия направлена на поддержание устойчивости водных экосистем. Насколько можно судить по показателям качества вод и эффективности процессов самоочищения, они по-прежнему сохраняются на высоком экологическом уровне. Анализ таксономического состава сообществ фитопланктона и микрофитобентоса показывает, что среди цианобактерий распространены виды, характерные для загрязненных местообитаний. Повсеместно с оценками обилия «в массе» и «очень часто» в течение всех лет исследований наблюдаются Phormidium foveolarum (Mont.) Gom., Spirulina jenneri (Hass.) Kütz. В 2010–2014 гг. видовое разнообразие таких организмов увеличилось. Например, распространились также такие виды как Anabaena constricta (Szaf.) Geitl., A. solitaria Klebs, Aphanothece clathrata W. et G. S. West, Ostillatoria lauterbornii Schmidle, O. princeps, O. putrida Schmidle. С оценками обилия «часто» и «нередко» распространены Microcystis aeruginosa f. pseudofilamentosa (Crow) Elenk., M. ichtyoblabe Kütz., Aphanothece stagnina (Spreng.) B.-Peters. et Geitl., Phormidium molle (Kütz.) Gom., Ph. uncinatum (Ag.) Gom., Ostillatoria аgardhii Gom., O. simplicissima Gom., O. splendida Grev., O. tenius Ag. и другие. В оз. Кипец в заливах Мохов Угол и Кипец «в массе» развит вид Ostillatoria coerulensis Gicklh. Все названные таксоны характерны для β-мезосапробных и полисапробных вод. Среди диатомовых водорослей в водоемах заповедника повсеместно распространены таксоны, характерные для вод II и III классов, по разряду качества относимых к вполне чистым, достаточно чистым и слабо загрязненным (Баринова и др., 2006). С высокими оценками обилия «нередко» и «часто» наблюдаются таксоны β-α-мезосапробных вод, такие как Fragilaria construens var. venter (Ehr.) Grun., Synedra parasitica (W. Sm.) Hust., Navicula cincta (Ehr.) Kütz., N. cuspidata Kütz., N. hungarica var. capitata Cl., N. lanceolata (Ag.) Kütz. et var. tenuirostris Skv., N. menisculus Schum., N. viridula Kütz., Anomoeoneis sphaerophora (Kütz.) Pfitz., Caloneis amphisbaena (Bory) Cl., Gomphonema angustatum var. productum Grun., Nitzschia recta Hantzsch., N. sigma (Kütz.) W. Sm., N. sublinearis Hust., N. triblionella var. debilis (Arn.) A. Mayer и таксоны α-β-мезосапробных вод, такие как Cyclotella meneghiniana Kütz., Gomphonema ventricosum Greg., Cymatopleura solea (Bréb.) W. Sm., а также α-мезосапробы, такие как Synedra tabulata (Ag.) Kütz., Navicula cryptocephala Kütz. et var. veneta (Kütz.) Grun., N. rhynchocephala Kütz., Nitzschia acicularis W. Sm., N. angustata (W. Sm.) Grun., N. apiculata (Greg.) Grun., N. hungarica Grun., N. triblionella Hantzsch., Hantzschia amphioxys (Ehr.) Grun. и другие.
Выводы Для сообществ фитопланктона и микрофитобентоса, изученных в 2007–2014 гг. в озерах Рамза и Кипец, характерно высокое видовое богатство (Анциферова, Богатырева, 2009; Анциферова, Борисова, 2009). Озерные экосистемы среднего течения р. Ворона представляют собой рефугиумы, в которых формируется и сохраняется видовое разнообразие диатомовых водорослей и цианобактерий, характерных для природных водных экосистем средних широт умеренного географического пояса. Они выступают как экологические коридоры, которые при возникновении благоприятных экологических условий могут способствовать их распространению в водоемах региона. Сложившаяся ситуация подтверждает (наиболее наглядно на примере цианобактерий), что данные водоемы являются открытыми экосистемами, в которых происходит исчезновение одних видов, вселение и распространение других, в соответствии со сложившимися климатическими и экологическими условиями (Анциферова, 2013; Толмачев, 1974). На основе изучения сообществ диатомовых водорослей и цианобактерий проточно-русловых озер заповедника в 2007–2009 гг. установлена развитая способность саморегулирования водных экосистем. Это подтверждается материалами многолетнего контроля эколого-биологического качества вод (Русова, 2015б). Высокая способность саморегулирования определяется эластичностью водных экосистем на уровне, обеспечивающем их устойчивое функционирование (Механизм устойчивости геосистем, 1992). Структурные перестройки микросообществ являются мерой устойчивости водных экосистем, которая контролируется сохранением и/или нарушением их эколого-биологического качества. Аномально высокие температуры воздуха 2010–2012 гг. выявили влияние данного природного фактора на устойчивое функционирование водных экосистем. Начиная с 2010 г., начались структурные перестройки в сообществах фитопланктона и микрофитобентоса (в сопоставлении с 2007-2009 гг.), направленные на сохранение устойчивости озерных экосистем. Они фиксируются как по изменениям общего числа таксонов, так и на таксономическом и экологическом уровнях. Наиболее наглядно данный процесс прослеживается на примере сообществ цианобактерий (Анциферова, 2013). К 2012 году таксономическая структура микросообществ практически полностью перестроились, вплоть до массового вселения новых для региона видов. Но при этом качество вод сохранилось на высоком уровне 2007–2009 гг. Как показали исследования, проведенные в 2013–2014 гг., в связи с возвратом температурных условий к среднестатистическим для региона параметрам, реакция экосистем вновь проявилась в значительных структурных перестройках микросообществ. Но при этом на фоне изменений их таксономического и экологического состава, в водоемах сформировались и распространились зоны полисапробных вод. В целом качество вод сохранилось на высоком уровне 2007–2009 гг. Водные экосистемы вновь проявили свойство эластичности, и перешли в новый бассейн устойчивости. Режим особо охраняемой природной территории способствует сохранению экологического благополучия водных экосистем заповедника. Это позволяет выявить и проследить реакцию сообществ фитопланктона и микрофитобентоса на изменившиеся природные условия, которая направлена на поддержание устойчивости водных экосистем проточно-русловых озер. Проточно-русловые озера отличает многообразие местообитаний и высокая способность к самоочищению вод. Состояние кризисности водных экосистем свидетельствует об обратимости происходящих в них процессов, подчеркивает экологическую опасность антропогенного загрязнения в условиях современной эпохи климатических изменений, в том числе глобального потепления.
Список литературы 1. Акимов Л.М., Задорожная Т.Н. Исследование динамики многолетнего ряда температуры в целях разработки сверхдолгосрочного прогноза // Вестник Воронежского государственного университета. Серия География. Геоэкология. № 3. – Воронеж, 2014. – С. 35–39. 2. Акимов Л.М., Акимов Е.Л. Пространственное распределение метеорологических параметров, влияющих на жизнедеятельность человека на территории Центрального Черноземья // Вестник Воронежского государственного университета. Серия География. Геоэкология. № 2. – Воронеж, 2014. – С. 17–24. 3. Анциферова Г.А. Разнообразие биотопов и видовое богатство микрофитоценозов проточно-русловых озер р. Вороны // «Проблемы изучения краевых структур биоценозов»: материалы 3-й Междунар. науч. конф. – Саратов: Изд-во Сарат. ун-та, 2012. – С. 16–22. 4. Анциферова Г.А. Особенности вегетационных сукцессий низших водорослей в условиях аномально высоких летних температур 2010-2012 годов (бассейн Среднего Дона) // Вестник Воронежского университета. Серия: География. Геоэкология. № 2. – Воронеж, 2013. – С. 107–112. 5. Анциферова Г.А., Богатырева С.Н. Низшие синезеленые и диатомовые водоросли Воронинского государственного природного заповедника // Труды гос. прир. заповедника «Воронинский». Т. 1. – Тамбов: Изд-во Першина Р.В., 2009. – С. 52–106. 6. Анциферова Г.А., Борисова Л.Е. Озера долины реки Вороны как естественный современный рефугиум диатомовых водорослей центра Восточно-Европейской равнины. // Вестник Воронежского университета. Серия: География. Геоэкология, № 2. – Воронеж, 2009. – С. 85–92. 7. Баринова С.С., Медведева Л.А., Анисимова О.В. Биоразнообразие водорослей-индикаторов окружающей среды – Тель-Авив, 2006. – 496 с. 8. Летопись Природы ФГУ «Государственный природный заповедник “Воронинский”». Кн. 17. – Инжавино, 2013. – 93 с. 9. Механизм устойчивости геосистем. – М.: Наука, 1992. – 208 с. 10. Русова Н.И. Эколого-биологическое качество вод озер Рамза и Симерка в долине реки Вороны (государственный природный заповедник «Воронинский») // Вестник Воронежского государственного университета. Серия География. Геоэкология. № 1. – Воронеж, 2013. – С. 87–93. 11. Русова Н.И. Эколого-географический анализ по диатомовым водорослям среднего течения р. Ворона // Школа экологических перспектив: материалы четвертого молодежного инновационного проекта: Воронеж, 2015а. – С. 130–132. 12. Русова Н.И. Оценка экологического состояния вод среднего течения р. Ворона на основе биоиндикационных исследований по сообществам низших водорослей // Современные проблемы науки и образования. 2015б. №1. – https://www.science-education.ru/ru/article/view?id=18054 (дата обращения 10.12.2016) 13. Садчиков А.П., Кудряшов М.А. Экология прибрежно-водной растительности. – М.: Изд-во НИА-Природа, РЭФИА, 2004. – 220 с. 14. Толмачев А.И. Введение в географию растений: (Лекции, читанные студ. Ленинградского ун-та в 1958-1971 гг.). – Л.: Изд-во Ленинградского ун-та, 1974. – 244 с. Статья поступила в редакцию 19.12.2016
Changes of stucture of microcommunities as a criterion of stability of natural aquatic ecosystems Galina A. Antsiferova, Nadezhda I. Rusova Lake Kipets and lake Ramza are located in the middle flow of the Vorona River. The structure of microcommunities represented by diatoms and cyanobacteria was studied during several last years. Changes of taxonomic structure of phytoplankton and phytobenthos to be considered as a criterion of stability of aquatic ecosystems. A bioindicative method was used for monitoring of ecosystem functioning and environmental safety. Key words: bioindication; aquatic ecosystem; diatoms; microalgae community; stability of aquatic ecosystem; functioning of aquatic ecosystem; cyanobacteria; ecology of aquatic ecosystem.
Об авторах Анциферова Галина Аркадьевна – Antsiferova Galina A. доктор географических наук g_antsiferova@mail.ru Русова Надежда Ивановна - Rusova Nadezhda I. преподаватель, ВМПИ ВУНЦ ВМФ «Военно-морская академия» им. Н.Г.Кузнецова, г. Санкт-Петербург, Россия (Navy VMPI VUNTS Navy «Naval Academy», St.-Petersburg, Russia), факультет турбинных энергетических установок, применения и эксплуатации вооружения средств радиационной, химической и биологической защиты сил флота, кафедра радиационной, химической и биологической защиты nadezhda_minnikova@mail.ru Корреспондентский адрес: 394068, Россия, г. Воронеж, ул. Хользунова, д. 40, корп. 5, Воронежский государственный университет.
ССЫЛКА НА СТАТЬЮ: Анциферова Г.А., Русова Н.И. Структурные перестройки микросообществ как мера устойчивости природных водных экосистем // Вопросы современной альгологии. 2017. № 1 (13). http://algology.ru/1095 Уважаемые коллеги! Если Вы хотите получить версию статьи в формате PDF, пожалуйста, напишите в редакцию, и мы ее вам с удовольствием пришлем бесплатно.
При перепечатке ссылка на сайт обязательна
К разделу ОБЗОРЫ, СТАТЬИ И КРАТКИЕ СООБЩЕНИЯ

|
|||
|
| ||